


Таблица 1 — Праймеры, используемые для выявления различных регионов сag A гена H. pylori
Сиквенс праймеров 5/–3/ | Температура отжига, °С | Ссылка | Размер фрагмента |
GATAACAGGCAAGCTTTTGAGGGA CTGCAAAAGATTGTTTGGCAGA | 55 | [11] | 349 п. н. |
TAACGCTGTCGCTTCATACG CCATTGGTAССTGTAGAAAC | 55 | Подобрана автором | 867 п. н. |
TGTTTGGGAAATGGATTGGT TTGCCCGCATACAATAATGA | 52 | Подобрана автором | 712 п. н. |
GGCAATGGTGGTCCTGGAGCTAGGC GGAAATCTTTAATCTCAGTTCGG | 53 | [11] | 324 п. н. |
GGAACCCTAGTCGGTAATG TTTCAAAGGGAAAGGTCCGCC | 53 | [12] | 501 п. н. |
GGAACCCTAGTCGGTAATG AGAGGGAAGCCTGCTTGATT | 53 | [12] | 495 п. н. |
В результате проведенного исследования было установлено, что фрагмент сagA-гена, характеризующий восточный тип (искомый фрагмент 501 п. н.), не был выявлен ни в одном из исследуемых образцов, несмотря на отработку условий амплификации. Данный факт свидетельствует о принадлежности циркулирующих в белорусском регионе штаммов H. pylori к западному типу. Так как сagA-ген является цитотоксинассоциированным и его присутствие определяет активность воспаления, нами проведена оценка соответствия выявленных сagA-генотипов и данных гистологического метода диагностики, а также сравнение чувствительности и специфичности 5 различных пар праймеров, используемых для выявления сagA-генотипа. Результаты представлены в таблице 2.
Как видно из таблицы 2, положительные результаты выявления сagA-генотипа, полученные всеми праймерами, составили 7 случаев при малоактивной форме, 7 — при умеренноактивной и 17 — при высокоактивной, то есть 31 случай выявления. При проведении анализа любой положительный результат учитывали как истинно положительный. Поэтому ложноположительные результаты приняты за 0, соответственно, и специфичность всех вариантов ПЦР (используемых праймеров) составила 100 %. Исходя из данных предпосылок сравнивали чувствительность праймеров для выявления CagA-генотипа при помощи программы «MedCalc».
Максимальная чувствительность получена для праймеров с фрагментом амплификации 349 п. н. Следует отметить, что данная последовательность праймеров наиболее часто используется исследователями при изучении распространенности сagA-позитивных штаммов как среди западной, так и среди восточной популяции H. рylori, несмотря на достаточно высокий полиморфизм сagA-гена.
На рисунке 1 представлены ROC-кривые чувствительности 5 используемых пар праймеров.
Таблица 2 — Гистологические показатели СОЖ, cвязанные с сagA-генотипами H. pylori у пациентов с хроническим гастритом в зависимости от используемых праймеров
Степень гастрита / статистические показатели | Размер амплифицируемого фрагмента гена сagA (пар нуклеотидов) | ||||
349 п. н. | 867 п. н. | 712 п. н. | 400 п. н. | 495 п. н.. | |
Частота выявления искомого фрагмента гена CagA | |||||
Малоактивный | 7 (38,89 %) n = 18 | 2 (11,11 %) n = 18 | 1 (5,56 %) n = 18 | 5 (27,78 %) n = 18 | 5 (27,78 %) n = 18 |
Умеренноактивный | 5 (45,45 %) n = 11 | 5 (45,45 %) n = 11 | 0 (0 %) n = 11 | 6 (54,55 %) n = 11 | 7 (63,64 %) n = 11 |
Высокоактивный | 17 (73,91 %) n = 23 | 5 (21,74 %) n = 23 | 3 (13,04 %) n = 23 | 6 (26,09 %) n = 23 | 13 (56,52 %) n = 23 |
Всего | 29 (55,7 %) n = 52 | 12 (23,07 %) n = 52 | 4 (7,69 %) n = 52 | 17 (32,69 %) n = 52 | 25 (48,07 %) n = 52 |
Чувствительность, % | 93,55 | 38,71 | 12,9 | 54,84 | 80,65 |
95 % доверительный интервал | 78,54–99,02 | 21,87–57,81 | 3,7–9,85 | 36,04–72,7 | 62,52–92,5 |
Достоверность различий | P1–2 < 0,001 P1–3 < 0,001* P1–4 = 0,003* P1–5 = 0,141 | P2–4 = 0,343 P2–3 = 0,2 | P3–5 < 0,001* P3–4 = 0,039* | P4–5 = 0,076 | P2–5 = 0,008* |
* Различия достоверны (р < 0,05)
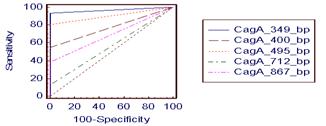
Рисунок 1 — ROC-кривые чувствительности используемых пар праймеров
для выявления фрагмента сagA-гена H. pylori
При отработке условий проведения ПЦР по определению исследуемых нами генотипов и аллельных вариантов H. рylori использовали те же препараты ДНК, методику и принцип подбора праймеров, что и при отработке условий амплификации cagA-генотипа. Подобранные праймеры для амплифицируемых регионов H. pylori представлены в таблице 3.
Таблица 3 — Последовательности праймеров и режим амплификации для выявления генотипов и аллельных вариантов H. pylori
Амплифицируемый регион H. pylori | Сиквенс праймеров 5/–3/ | Температура отжига, °С | Ссылка | Размер фрагмента |
сagA | GATAACAGGCAAGCTTTTGAG GGA CTG CAAAAGATTGTT TGGCAGA | 55 | [11] | 349 п. н. |
vacAs (s1 and s2) | ATGGAAATACAACAAACACAC CTGСTTGAATGCGCCAAAC | 55 | [1] | 286 п. н. |
vacAm1a | GGTCAAAATGCGGTCATGG CCATTGGTAССTGTAGAAAC | 55 | [1] | 290 п. н. |
vacAm1b | GGCCCCAATGCAGTCATGGAT GCTGTTAGTGCCTAAAGAAGCAT | 57 | [1] | 291 п. н. |
vacAm2 | GGAGCCCCAGGAAAC ATTG CATAACTAGCGCCTTGCAC | 58 | [1] | 352 п. н |
vacAs1a | GTCAGCATCACACCGCAAC ATGGAAATACAACAAACACAC | 50 | [1] | 190 п. н. |
vacAs1b | GTCAGCATCACACCGCAAC ATGGAAATACAACAAACACAC | 50 | [1] | 187 п. н. |
vacAs1c | CTYGCTTTAGTRGGGYTA ATGGAAATACAACAAACACAC | 50 | [1] | 213 п. н. |
iceA1 | GTGTTTTTAACCAAAGTATС CTATAGCCASTYTCTTTGCA | 43 | [13] | 247 п. н. |
iceA2 | GTTGGGTATATCACAATTTAT TTRCCCTATTTTCTAGTAGGT | 46 | [13] | 229–334 п. н. |
babA2 | AATCCAAAAAGGAGAAAAAGTATGAAA TGTTAGTGATTTCGGTGTAGGACA | 58 | [14] | 800 п. н. |
jhp 0917 (plasticity regions) | TGGTTTCTACTGACAGAGCGC AACACGCTGACAGGACAATCTCCC | 50 | [15] | 307 п. н. |
jhp 0918 (plasticity regions) | CCTATATCGCTAACGCGCGCTC AAGCTGAAGCGTTTGTAACG | 55 | [15] | 276 п. н. |
Для проведения ПЦР применялись однотипные условия амплификации, отличающиеся только температурой отжига, которая для каждой пары указана в таблице 1 и 3: 95 °C, 5 минут; 30 × (95 °C, 30 секунд; 55 °C, 30 сеунд; 72 °C, 1 минута); 72° C, 1 минута.
Распределение генотипов и аллельных вариантов H. pylori — саgA, vacAs (s1 and s2), vacA m1a, vacAm1b, vacAs1a, vacA m2, vacAs1b, vacAs1c, bab A2, iсeA1, iсeA2, jhp 0917, jhp 0918 в исследуемой группе пациентов с ХГ (n = 52) представлены в таблице 4.
Таблица 4 — Распределение генотипов и аллельных вариантов H. pylori в группе пациентов с диагнозом хронический гастрит
Cубтипы H. pylori | Количество выявленных случаев | Частота, % |
сag A | 29 | 55,7 |
vacAm | — | — |
vacAm1a | 20 | 38,5 |
vacAm1b | 5 | 9,6 |
vacAm2 | 23 | 44,2 |
vacAs | — | — |
vacAs1a | 23 | 44,2 |
vacAs1b | 1 | 1,9 |
vacAs1c | 0 | 0 |
vacAs2 | 23 | 44,2 |
babA2 | 1 | 1,9 |
ice A1 | 1 | 1,9 |
ice A2 | 2 | 3,8 |
jHP 0917 | 1 | 1,9 |
jHP 0918 | 0 | 0 |
cag A + vacAm1/s1 | 14 | 26,9 |
cag A + vacAm1/s1 + ice A1 + babA2 | 1 | 1,9 |
cag A + vacAm1/s1 + ice A2 | 2 | 3,8 |
cag A + vacAm1/s1 + vacAm2 / vacAs1 | 8 | 15,4 |
ИТОГО микст-штаммы | 25 | 48,1 |
Проведенные нами исследования показали, что практически в половине случаев — 48,1 % среди пациентов с ХГ присутствуют микст-штаммы c несколькими патогенными генотипами, в том числе в 15,4 % микст-штаммы с различными аллельными вариантами вакуолизирующего цитотоксина vacAm1 + vacAm2 / vacAs1 + vacAs2.
Разработанная нами методика позволила провести оценку генотипического разнообразия H. pylori и роли микст-штаммов с различными аллельными вариантами в формировании патологии желудочно-кишечного тракта в выборочной группе пациентов с ЯБДК, ЯБЖ, НПВП-гастропатиями в сравнении с группой ХГ.
Результаты проведенного исследования представлены в таблице 5.
В результате данного исследования по большинству выявляемых генотипов определена сходная генетическая структура штаммов H. pylori при ЯБДК, ЯБЖ, НПВП-гастропатиях и ХГ, что может быть связано с региональными особенностями субтипов H. pylori. Тем не менее генотип babA2, ice A1, ice A2 достоверно реже встречался в группе пациентов с ХГ по сравнению с группами больных ЯБДК и НПВП-гастропатиями, генотипы vacAm1, vacAm2, vacAs1, vacAm1s1+ vacAm2s2, ice A2 достоверно реже встречались в группе с гастритами по сравнению с группой с ЯБДК (р < 0,05). Полученные нами данные свидетельствуют о том, что менее агрессивные штаммы способствуют развитию гастритов и не приводят к развитию язв.
Oценку значимости саg A-генотипа в формировании патологии ЖКТ в белорусском регионе проводили в группах пациентов c заболеваниями желудочно-кишечного тракта (n = 226) в сравнении с контрольной группой (n =24) и группой больных ХГ (n = 91) (таблица 6).
При анализе выявления сag A-генотипа H. pylori в контрольной и сравниваемых группах показано, что выявлены достоверные различия при сравнении контрольной группы и группы пациентов с ЯБДК (p = 0,007). Сag A-генотип H. pylori выявлен в 83 % исследуемых образцов. Проведенные нами исследования показывают значительный вклад цитотоксинассоциированного саgA-гена H. pylori в формирование ЯБДК в белорусском регионе.
Таблица 5 — Распределение генотипов H. pylori в группах обследованных пациентов с заболеваниями ЖКТ
Субтипы H. pylori | Группы обследованных | Достоверность различий | ||||||||
1. ЯБДК, n = 68 | 2. ЯЖ, n = 12 | 3. НПВП-гастропатии, n = 16 | 4. ХГ, n = 52 | χ2 | р | |||||
n | % | n | % | n | % | n | % | |||
cagA | 46 | 67,6 | 9 | 75 | 6 | 37,5 | 29 | 55,7 | 1,77 0,8 0,03 | p1–4 = 0,18 p2–4 = 0,37 p3–4 = 0,85 |
babA2 | 30 | 44,1 | 3 | 25 | 3 | 18,8 | 1 | 1,9 | 25,2 3,59 5,36 | p1–4 <0,01* p2–4=0,58 p3–4=0,02* |
vacAm1 | 46 | 67,6 | 10 | 83,3 | 6 | 37,5 | 25 | 48,1 | 4,67 3,57 0,21 | p1–4=0,03* p2–4=0,06 p3–4=0,65 |
vacAm2 | 51 | 75 | 8 | 66,7 | 6 | 37,5 | 23 | 44,2 | 11,8 1,17 0,03 | p1–4 = 0,0006* p2–4=0,27 p3–4=0,85 |
vacAs1 | 50 | 73,5 | 9 | 75 | 6 | 37,5 | 24 | 46,1 | 9,34 2,2 0,1 | p1–4=0,002* p2–4=0,13 p3–4=0,74 |
vacAs2 | 37 | 54,4 | 7 | 58,3 | 6 | 37,5 | 23 | 44,2 | 1,22 0,32 0,03 | p1–4=0,26 p2–4=0,57 p3–4=0,85 |
vacAm1s1+ vacAm2s2 | 22 | 32,4 | 4 | 33,3 | 4 | 25 | 8 | 15,4 | 4,52 1,05 0,26 | p1–4=0,03* p2–4=0,3 p3–4=0,6 |
ice A1 | 10 | 14,7 | 0 | 0 | 6 | 75 | 1 | 1,9 | 5,78 13,1 | p1–4=0,01* p3–4=0,0003* |
ice A2 | 23 | 33,8 | 3 | 25 | 5 | 62,5 | 2 | 3,8 | 16,1 3,48 7,2 | p1–4=0,0001* p2–4=0,06 p3–4=0,0073* |
jHP0917+ jHP0918 | 5 | 7,35 | 1 | 8,3 | 0 | 0 | 0 | 0 |
Поимечание. * Различия достоверны (р < 0,05); n — количество обследованных пациентов в группе; n — количество обследованных пациентов с положительным субтипом.
Таблица 6 — Частота выявления сagA генотипа H. рylori среди пациентов с заболеваниями желудочно-кишечного тракта и контрольной группы
Группы (возрастной диапазон) | Выявлен сagA-генотип H. рylori | В сравнении с контрольной группой | В сравнении с группой больных гастритом | ||
χ2 | р | χ2 | р | ||
Контрольная группа, n = 24 | 12 (50 %) | ||||
ХГ, n = 91 | 62 (68,13 %) | 2,72 | P 1–2 = 0,099 | ||
РЖ, n = 22 | 14 (63,64 %) | 0,4 | P 1–3 = 0,53 | 0,02 | P 2–3 = 0,88 |
ЯБЖ, n = 37 | 24 (64,86 %) | 1,33 | P 1–4 = 0,24 | 0,13 | P 2–4 = 0,72 |
ЯБДК, n = 48 | 40 (83,33 %) | 7,28 | P 1–5 = 0,007* | 2,98 | P 2–5 = 0,084 |
НПВП-гастропатии, n = 28 | 17 (60,71 %) | 0,6 | P 1–6 = 0,53 | 0,00 | P 2–6 = 0,46 |
Заболевания желудочно-кишечного тракта, n = 226 (взрослые) | ,49 %) | 2,26 | P 1–7 = 0,13 | ||
Заболевания желудочно-кишечного тракта без гастритов, n = 179 | ,42 %) | 1,62 | P 2-8 = 0,2 |
* При сравнении с контрольной группой различия достоверны (р < 0,05)
Заключение
Методика определения генотипов и аллельных вариантов H. pylori с использованием ПЦР позволяет определять генетическую структуру штаммов и их роль в развитии заболеваний ЖКТ. Достоверных различий между группами больных с заболеваниями ЖКТ и группой больных ХГ не выявлено (p > 0,05), что свидетельствует о близкой генетической структуре H. pylori в группах пациентов с заболеваниями ЖКТ и характеризует региональные особенности штаммов. Достоверно реже встречаемые при ХГ генотипы babA2, ice A1, iceA2 в сравнении с ЯБДК и НПВП-гастропатиями, vacAm1, vacAm2, vacAs1, vacAm1s1+ vacAm2s2 в сравнении с ЯБДК (р < 0,05) свидетельствуют о позитивном влиянии менее агрессивных генотипов на течение заболевания. Достоверно чаще встречается сagA-генотип при сравнении контрольной группы и группы пациентов с ЯБДК (p = 0,007), что показывает значимость данного гена в развитии дуоденальных язв в Республике Беларусь.
БИБЛИОГРАФИЧЕСКИЙ СПИСОК
1. Mosaicism in vacuolating cytotoxin alleles of Helicobacter pylori. Association of specific vacA types with cytotoxin production and peptic ulceration/ J. Atherton [et al.] // J Biol Chem. — 1995. — № 28. — P. 17771–17777.
2. The N-terminal 34 kDa fragment of Helicobacter pylori vacuolating cytotoxin targets mitochondria and induces cytochrome С release / A. Galmiche [et al.] // EMBO J. — 2000. — № 19. — Р. 6361–6370.
3. Covacci, A. Tyrosine-phosphorylated bacterial proteins: Trojan horses for the host cell / A. Covacci, R. Rappuoli // J. Ex. Med. — 2002. — № 000. — Р. 587–592/
4. Пиманов, С. И. Эзофагит, гастрит и язвенная болезнь: рук-во для врачей / . — Минск: Медицинская книга, 2000. — 378 с.
5. Современные представления об этиологии и патогенезе язвенной болезни / [и др.] // Вестник АМН СССР. — 1990. — № 3. — С. 60–64.
6. Helicobacter pylori adhesin binding fucosylated histo-blood group antigens revealed by retagging / D. Ilver [et al.] // Science.-1998. — № 000. — Р. 373–377.
7. Expression of Lewis b blood group antigen in Helicobacter pylori does not interfere with bacterial adhesion property / Peng-Yuan Zheng [et al.] // World J Gastroenterol. — 2003. — Vol. 9, № 1. — P.122-124.
8. Клиническое значение факторов патогенности Helicobacter pylori / [и др.] // Рос. журн. гастроэнтерол., гепатол., колопроктол. — 2005. — Т. 15, № 3. — С. 22–27.
9. Analysis of iceA1 transcription in Helicobacter pylori / J. P. Donahue [et al.] // Helicobacter. — 2000. — Vol. 15, № 1. —P. 1–12.
10. DupA as a risk determinant in Helicobacter pylori infection / M. Douraghi [et al.] // J. Med. Microbiol. — 2008. — Vol. 57. — P. 554–556.
11. Tummuru, M. Cloning and expression of a high molecular mass major antigen of Helicobacter pylori: Evidence of linkage to cytotoxin production / M. Tummuru, T. Cover, M. Blaser // Infect Immun. — 1993. — Vol. 61. — P. 1799–1809.
12. Argent, R. H. Simple method for determination of the number of Helicobacter pylori CagA variable-region EPIYA tyrosine phosphorylation motifs by PCR / R. H. Argent, Y. Zhang, J. C. Atherton // J. Clin. Microbiol. — 2005. — Vol. 43. — P. 791–795.
13. Adherence to gastric epithelial cells induces expression of a Helicobacter pylori gene, iceA, that is associated with clinical outcome / R. Peek [et al.] // Proc. Assoc Am Physicians. — 1998. — Vol. — P. 531–544.
14. Correlation of the Helicobacter pylori Virulence and Adherence Factors vacA, cagA and babA with Ulcer Disease in Four Different European Countries / M. Gerhard [et al.] // Gut. — 2001. — № 1.
15. Duodenal Ulcer Promouting Gene of Helicobacter pylori / H. Lu [et al.]// Gastroenterology. — 2005. — Vol. 128. — P. 833–848.
Поступила 29.03.2011
УДК 577.3+57.052
КОРРЕЛЯЦИЯ СТРУКТУРЫ И АНТИОКСИДАНТНОЙ АКТИВНОСТИ МОЛЕКУЛ
МЕЛАТОНИНА И ТРИПТОФАНА
, ,
Институт фармакологии и биохимии НАН Беларуси, г. Минск
Гродненский государственный аграрный университет
Гродненский государственный медицинский университет
Изучали радикал-скэвенджерные свойства мелатонина и триптофана в системах, генерирующих алкоксильные и пероксильные радикалы (эритроциты человека, обработанные окислителем терт-бутилгидропероксид (тБГП)). Структурные и электронные свойства мелатонина, триптофана и окисленных производных данных молекул были исследованы теоретически с использованием АМ1 полуэмпирического метода и ab initio метода. Высокая радикал-скэвенджерная способность мелатонина может быть обусловлена высокими значениями площади поверхности и дипольного момента молекулы, который отражает плотность заряда молекулы. С термодинамической точки зрения N1-ацетил - N2 -формил-5-метоксикинурамин (AФMK) является самым стабильным продуктом окисления мелатонина. Сравнительное исследование радикал-скэвенджерных, структурных и электронных свойства молекул мелатонина, его предшественника — триптофана продемонстрировало, что мелатонин в отличие от триптофана является потенциальным липидным антиоксидантом и эффективным скэвенджером алкоксильных и пероксильных радикалов.
Ключевые слова: структура, электронные свойства, радикал-скэвенжер, мелатонин, триптофан.
STRUCTURE AND ANTIOXIDANT WCTIVITY ASSOCIWTIONS
FOR MELATONIN MOLECULE AND ITS DERIVATIVTS
V. T. Cheshchevik, S. N. Sakolouskaya, E. A. Lapshina, S. N. Iljina
Instutute for Pharmacology and Biochemistry NAS Belarus, Minsk
Grodno State Agricultural University
Grodno State Medical University
Structural and electronic properties of melatonin, tryptophan, and oxidative products of these molecules were determined theoretically by performing semi-empirical and ab initio calculations. The radical scavenging effects of melatonin and tryptophan were studied using the alkoxyl and peroxyl radical - generating systems. We have shown that melatonin, rather than tryptophan, is a potent lipid antioxidant and an effective scavenger of alkoxyl - and peroxyl radicals.
Key word: structure, electronic properties, radical-scavenger, melatonin, tryptophan.
Введение
В ряде исследований продемонстрировано, что мелатонин является эффективным антиоксидантом и скэвенджером свободных радикалов [1]. Показано, что мелатонин связывает гидроксильные радикалы, токсические пероксильные радикалы, но, с другой стороны, не взаимодействует непосредственно с супероксиданион радикалом [2]. Ранее мы продемонстрировали, что мелатонин является эффективным скэвенджером гипохлорной кислоты [3]. С целью выяснения корреляций структуры молекулы мелатонина, его физико-химических свойств и биохимической активности нами было проведено сравнительное исследование радикал-скэвенджерных, структурных и электронных свойства молекул мелатонина, его предшественника — триптофана и их производных, определены возможные молекулярные центры связывания радикалов. Радикал-скэвенджерные свойства мелатонина и триптофана изучали в системах, генерирующих алкоксильные и пероксильные радикалы (эритроциты человека, обработанные окислителем тБГП). Структурные и электронные свойства мелатонина, триптофана и окисленных производных данных молекул были исследованы теоретически с использованием полуэмпирических (АМ1) методов и ab initio расчетов.
Материалы и методы исследований
В работе использовали бутилированный гидрокситолуол (БГТ), 2-тиобарбитуровую кислоту (ТБК), трихлоруксусную кислоту (ТХУ), додецил сульфат натрия, этилендиаминтетраацетат (ЭДTА), терт-бутилгидропероксид, 2, 2´- азобис (2-амидинпропан) дигидрохлорид (AAPH) (Sigma — Aldrich Chemie GmbH, Германия). Флуоресцентный зонд 1-(-триметиламмоний фенил)-6-фенил-1,3,5-гексатриен-р-толуолсульфонат (ТМА — DPН) получен от Molecular probes, Eugene, OR, USA. Все остальные реактивы производства «Реахим» (Россия). Все растворы готовили на воде, очищенной с помощью Milli-Q системы (Millipore), имеющей проводимость 18,4 MW cм–2.
Эритроциты. Кровь здоровых доноров (с добавлением 3 % раствора цитрата натрия) получали на Станции переливания крови г. Гродно. После удаления плазмы и слоя лейкоцитов эритроциты промывали 3 раза холодным (4 °С) изотоническим раствором NaCl, содержащим фосфатный буфер (ФБ: 0,145 M NaCl, 1,9 мM NaH2PO4 и 8,1 мM Na2HPO4, pH = 7,4). Эритроциты использовали немедленно после получения. Мембраны эритроцитов получали по методу Доджа и соавт. [4, 5]. Гемолизат готовили лизисом 0,1 мл суспензии эритроцитов (5 % гематокрит) в 1 мл H2O.
Перекисное окисление липидов определяли как количество ТБК-реактивных соединений (ТБКРС) в кислоторастворимой фракции суспензии эритроцитов по методу Стокса и Дормэнди [6].
Окислительный стресс индуцировали, инкубируя эритроциты человека (гематокрит 10 %) в ФБ c различными концентрациями тБГП при 21 °С в течение часа.
Результаты исследования и их обсуждение
На рисунке 1 показана концентрационная зависимость ингибирования мелатонином процессов перекисного окисления липидов (ПОЛ) в суспензии эритроцитов, обработанной тБГП. Ингибирование, вероятно, осуществляется прямым скэвенджированием алкоксильных и пероксильных радикалов, образуемых при разложении перекиси в эритроцитах. Другая возможность заключается во взаимодействии липидных пероксильных радикалов с мелатонином, что приводит к ингибированию процессов перекисного окисления мембранных липидов. Мы сравнили эффективность взаимодействия мелатонина, триптофана и известного липидного антиоксиданта БГТ со свободными радикалами и продемонстрировали ингибирование образования ТБКРС в суспензии эритроцитов (рисунок 1) или тушение люминол-зависимой хемилюминесценции данными эффекторами в системе генерации свободных радикалов Fe2+ — тБГП (рисунок 2).
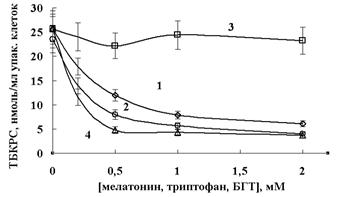
Рисунок 1 — Протекторный эффект мелатонина (1, 2), триптофана (3) или БГТ (4)
при тБГП-индуцированном перекисном окислении мембранных липидов в эритроцитах (1, 3, 4)
и в гемолизатах (2) того же количества клеток. Суспензию клеток (5% гематокрит) или гемолизат
в ФБ (рН 7,4) обрабатывали 1,25 мМ тБГП при 37 °С в течение 30 мин. Предварительно клетки
или гемолизат преинкубировали с различными концентрациями мелатонина,
триптофана и БГТ при 37 °С в течение 20 мин
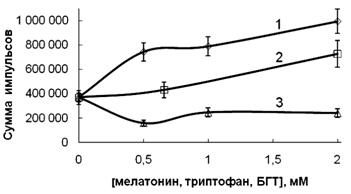
Рисунок 2 — Эффект мелатонина (1), триптофана (2) или БГТ (3) на люминол-зависимую
хемилюминесценцию, генерируемую в системе Fe2+ (1 мМ) + тБГТ (20 мМ) в ФБ (рН 7,4) при 37 °С
Концентрации мелатонина и липофильного антиоксиданта БГТ, вызывающие 50 % ингибирование окислительного процесса (IC50), составили 450 ± 55 и 180 ± 15 мкМ соответственно (рисунок 1). В то же время мелатонин не оказывал эффекта на внутриклеточные окислительные процессы: окисление внутриклеточного глутатиона и образование метНв. Как видно на рисунке 1, протекторный эффект мелатонина, наблюдаемый в гемолизатах эритроцитов после обработки тБГТ, был выше, чем в суспензии эритроцитов. На рисунке 2 показаны эффекты мелатонина, триптофана и БГТ на общую интенсивность хемилюминесценции, генерируемой в системе Fe2+ + тБГТ эритроцитах. Значения IC50 ингибирования генерации радикалов в системе «эритроциты/тБГТ» составили 490 ± 65 и 95 ± 11 мкМ для мелатонина и БГТ соответственно. Интересно, что мелатонин и в меньшей степени — триптофан повышал интенсивность люминол-зависимой хемилюминесценции, генерируемой в системе Fe2+ + тБГТ (рисунок 2).
Процессы тушения люминол-зависимой хемилюминесценции, генерируемой свободными радикалами, являются сложными, поэтому графики Штерна-Фольмера тушения хемилюминесценции не являются линейными. Используя графики Штерна-Фольмера для ингибирования образования TБКРС в эритроцитах, мы рассчитали кажущуюся константу Штерна-Фольмера, характеризующую взаимодействие мелатонина со свободными радикалами. Кажущаяся константа Штерна-Фольмера для мелатонина, которую определили из линейной части графика, составила (0,23 ± 0,05) × 104 М-1. Значения кажущихся констант Штерна-Фольмера для ингибирования перекисного окисления липидов в эритроцитах ВНТ и триптофаном, определенные из начальных линейных участков графиков Штерна-Фольмера, составили (0,85 ± 0,10) × 104 М-1 и (0,02 ± 0,005) × 104 М-1 соответственно.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 |
