

Резюмируя, мы считаем возможным происхождение полета из полета над экраном и глиссирования при спасении от опасности некрупных оперенных форм, обитающих на стыке биотопов прибрежных зарослей и открытых мелководий. При этом сочетании у форм, обнаруживающих добычу с помощью зрения, могли развиться приспособления, расширяющие отбрасываемую тень за счет удлинения перьев на передних конечностях при сохранении способности к бегу и лазанию в зарослях. Во всяком случае, эта гипотеза объясняет наличие соответствующих морфологических приспособлений при невозможности их использования в современной привычной роли, а также первичность для птиц активного машущего полета.
Взаимосвязь признаков в период становления класса птиц
В качестве основных признаков, характеризующих класс птиц, обычно приводят: 1) полет, 2) пе-рьевой покров, 3) высокий уровень метаболизма и гомойотермность при высокой температуре тела, 4) усиление заботы о потомстве, включающее насиживание, вождение и кормление потомства. Реже, но упоминают высокий уровень высшей нервной деятельности и прогрессивное развитие головного мозга (признак 5). Учитывая, что перьевой покров обеспечивает полет и сохранение температуры тела при минимальных дополнительных затратах энергии, можно было бы и не выделять его в качестве самостоятельного признака, но он удобен чисто морфологически. Мы предлагаем, особенно при обсуждении отличий ранних птиц от развитых рептилий, учитывать и другие признаки, характеризующие класс птиц: 6) детерминированные некрупные размеры взрослой формы, 7) увеличение размеров яйца, 8) уменьшение числа яиц в кладке, 9) неодновременное созревание яиц, 10) ускорение темпов роста при быстром достижении взрослых размеров.
Эти признаки образовывали при дифференциации птиц как класса достаточно жесткую систему со значительным числом обратных связей, благодаря которым у птиц выработались признаки, которые мы сейчас считаем типично птичьими, хотя, возможно, ряд из них присутствовал у близких по происхождению групп динозавров.
Рассмотрим взаимодействие этих признаков-адаптаций в процессе становления птиц как отдельной группы. Высокий уровень метаболизма и теплокровность при высокой температуре тела позволяли переходить к полету, который, в свою очередь, требовал этих признаков. Полет также требовал мелких размеров формы, прибегающей к нему, особенно при еще слабом развитии летательной мускулатуры и, в свою очередь, только некрупные размеры позволяли летать хотя бы изредка и совершенствовать способы полета. Но детерминированные некрупные размеры при значительном прессе хищников требовали увеличения выживания потомства. Это могло быть достигнуто двумя способами: при r-стратегии увеличением плодовитости, т. е. в нашем случае увеличением количества откладываемых яиц. Ряд современных форм птиц прибегает к этому методу (куриные, утки, ряд воробьиных (Paridae, Sittidae)), но ранние птицы выходили из положения по пути К-стратегии, т. е. увеличением выживаемости небольшого числа потомков. Это требовало усиления заботы о потомстве, в том числе интенсификации насиживания, более тщательной заботы о выводке, вождения и кормления его. Усиление заботы о потомстве позволяло уменьшить число яиц до нескольких (по-видимому не более 2–4, как у некоторых современных гекконов, у которых к тому же сходное соотношение размеров яйца и взрослой особи). При этом, возможно, у птиц еще наблюдалось одновременное откладывание яиц одной кладки. В свою очередь, уменьшение числа яиц в кладке требовало усиления заботы о потомстве. Пресс наземных хищников наиболее существенно сказывался в период откладки яиц, насиживания и вождения нелетных птенцов. Это требовало сокращения гнездового периода, что могло быть обеспечено в первую очередь усилением темпов роста, и, возможно, более быстрым достижением окончательных размеров, пригодных для полета. Усиление темпов роста было возможно только при усилении заботы о потомстве, в первую очередь кормлении. Сокращение гнездового периода и сроков роста было возможно и при увеличении величины яиц, и, как следствие, стартового размера птенца. Однако, увеличение размеров яйца шло вразрез с требованиями полета взрослой формы, и птицы выработали механизм последовательной откладки яиц. При небольшой кладке это позволяло сохранить привязанность к месту кладки (гнезду) и в дальнейшем насиживанию. Неодновременное созревание яиц, таким образом, позволяло еще более увеличить размеры яйца, так как для формирования каждого следующего яйца использовались новые порции пищи, а не запасы питательных веществ, как у рептилий. Сложность синхронизации созревания крупных фолликулов в парных яичниках и, по-видимому, еще большая сложность синхрнонного формирования белковых и скорлуповых оболочек перед откладкой яиц заставила птиц отказаться сначала от парных яйцеводов, а потом и яичников. К чему приводит одновременное созревание двух яйцеклеток, хорошо видно у современных высокопродуктивных пород кур, где часто встречаются двухжелтковые яйца с практически нулевым выходом живых цыплят. Формы с одним яйцеводом и одним яичником, таким образом, получали преимущества только при асинхронном созревании и неодновременной откладке яиц. Неодновременная откладка яиц позволяла сохранять способность к полету взрослой формы и некрупные размеры взрослой формы, а также при уменьшенном количестве яиц в кладке, быстрому переходу от откладки яиц к насиживанию и вождению. Таким образом, все перечисленные выше признаки могли “работать” и быть адаптивными только в комплексе. Кроме того, усиление заботы о потомстве требовало повышения уровня высшей нервной деятельности для увеличения выживаемости как молодняка, так и взрослой формы. Причем это привело как к развитию сложных инстинктов, так и к способности к научению, сохранению и обработке информации взрослой особью и передачи ее молодняку. Взаимосвязь данных признаков см. на схеме (рис. 4). В продвинутых группах птиц за счет улучшения летных качеств и дальнейшего усиления заботы о потомстве жесткость связей между рассмотренными признаками ослабляется. При этом становятся возможными увеличение размеров взрослой формы, уменьшение числа яиц в кладке (до одного) и увеличение его (до 10–15), удлинение срока гнездового периода, удлинение и укорочение срока взросления и длительности жизни взрослой особи.
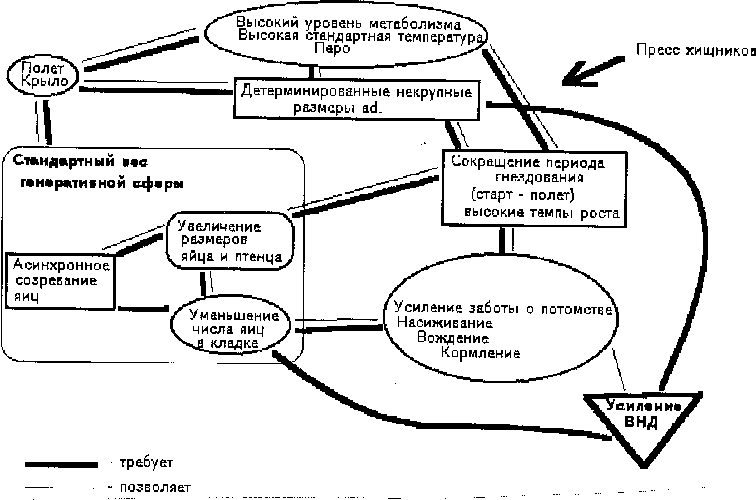
Рис. 4. Схема взаимосвязи признаков в период становления класса птиц.
В свете рассмотренной схемы связей обращает на себя внимание параллелизм в способах доведения потомства до жизнеспособных стадий у млекопитающих и птиц, а также некоторых рыб и рептилий, хотя решения сходных задач достигаются разными способами. У некоторых хрящевых и костных рыб, а также некоторых рептилий потомство развивается в организме матери до довольно крупных размеров, чаще только за счет крупного яйца, но иногда и за счет организма матери. У ряда этих животных наблюдается охрана выводка и вождение его до довольно крупных размеров. Очень редко, но наблюдается кормление выводка взрослой особью естественными кормами и у единичных видов за счет организма самки. Ни у кого из этих групп данные приспособления не присутствуют в комплексе. Передача потомству благоприобретенных форм поведения практически отсутствует.
У млекопитающих эмбрионы развиваются в организме матери при высокой постоянной температуре за счет организма матери. Для большинства млекопитающих характерно согревание выводка и вождение его до достижения достаточно крупных размеров. Наблюдается кормление выводка естественными кормами и всегда питание на ранних этапах за счет организма матери (молоком). Обычна высокая информационная связь и передача навыков потомству.
У птиц развитие происходит хотя и вне организма матери, но под ее контролем при создании ею высокой константной температуры. Условия в гнезде под насиживающей птицей сходны с внутриутробными, хотя развитие эмбриона идет за счет крупного яйца. Характерны охрана и вождение потомства фактически до достижения взрослых размеров и самостоятельности. Практически у всех птиц наблюдается кормление специфическими естественными кормами, либо первое время за счет организма взрослой особи веществами, сходными по составу с молоком (голуби, пингвины, фламинго). Наблюдается высокая информационная связь и передача приобретенных навыков потомству.
Выводы
1. На формирование характерного для птиц таза, и в первую очередь разворота назад и в стороны лобковых и седалищных костей, влияло явление втягивания в брюшную полость птенца через широкий пупочный канал значительного объема желтка — необходимого запаса для повышения выживаемости в первые дни жизни у гомойотермных выводковых птиц. Этому способствовали крупные размеры птенца, некрупные — взрослой формы, малое время и быстрые темпы роста.
2. Развитие пера из чешуйчатого покрова предшествовало полету и первоначально его подвижность использовалась для демонстраций (зрительное увеличение объема тела), и только затем перьевой покров принял на себя функцию термоизоляции.
3. Местом формирования ранних форм птиц являлись прибрежные мелководья с сочетанием открытых мест и зарослей, что обусловило способность к двуногому бегу с сохранением способности к лазанию.
4. Развитие маховых перьев (еще непригодных для полета) стимулировалось использованием их для создания тени при визуализации добычи (типа современной охоты “под навесом” цапель).
5. Первым полетом был машущий (хлопающий) полет-глиссирование по поверхности воды, не требующий развитой несущей поверхности крыльев и особо сильных полетных мышц, использущийся для спасений от опасности в зарослях.
6. В силу выгодности, хотя и крайней кратковременности, полуполета-глиссирования при спасении от наземных хищников он получил дальнейшее развитие, причем сначала как полет над экраном, требующий меньших энергозатрат, а потом как транзитный полет — самый распространенный тип полета среди всех групп птиц.
7. В период становления птиц как класса возник ряд адаптаций организованных в систему с обратными связями и включающий как общепринятые: способность к полету, высокий уровень иетаболизма, усиление заботы о потомстве, так и считавшиеся менее существенными: некрупные детерминированные размеры взрослых, быстрые темпы роста молодых, асинхронность откладки очень небольшого количества относительно крупные яиц. Эта система позволяла и заставляла совершенствовать полет и повышать уровень высшей нервной деятельности как наиболее существенные адаптации, позволяющие снижать смертность при выраженной К-стратегии.
Список литературы
, , (1965): Определитель млекопитающих СССР. М.: Просвещение. 1-379.
(1981): Записки палеонтолога. Л.: Наука. 1-164.
(1986): Зоологические путешествия. Л.: Наука. 1-199.
, (1984): Анатомия и гистология домашней птицы. М.: Колос. 1-287.
(1949): Биологические основы полета птиц. М.: МОИП. 1-248.
(1991): Эволюционный процесс. Критический обзор эволюционной теории. М.: Мир. 1-486.
, (1988): Редкие и исчезающие животные. Земноводные и пресмыкающиеся. М.: Высшая школа. 1-463.
(1940): Руководство по зоологии. Птицы. М.-Л. 6: 1-856.
(1965): Систематика птиц. Современное состояние и некоторые проблемы. - Итоги науки. Зоология. М. 5-56.
(1975): Миграционное состояние птиц. М.: Наука. 1-397.
(1982): Методы изучения бюджетов времени и энергии у птиц. - Бюджеты времени и энергии у птиц в природе. Л.: АН СССР. 124-153.
. (1982): Продукция и продуктивная энергия при откладке яиц у птиц. - Там же: 3-36.
(1981): Поведение животных. Сравнительные аспекты. М.: Мир. 1-479.
(1975): Рептилии. М.: Мир. 1-190.
(1974): Систематика птиц. М.: Высшая школа. 1-368.
(1967): Полет над экраном и глиссирование птиц. - Вестн. зоол. 2: 3-8.
(1966): Морфология и поведение (на примере пищедобывательной активности цапель). - Механизмы полета и ориентации птиц. М.: Наука. 169-223.
(1966): Некоторые соображения о бионических исследованиях в отнитологии. - Механизмы полета и ориентации птиц. М.: Наука. 5-25.
(1970): Энергетика полета насекомых и птиц. - Ж. общ. биол. 5: 527-549.
(1982): История жизни. Киев: Н. думка. 1-217.
, , (1991): Террариум и его обитатели. М.: Лесн. пром-сть. 1-349.
(1987): Авимимиды и проблема происхождения птиц. М.: Наука. 1-93.
(1978): Рукокрылые охотники. М.: Лесн. пром-сть. 1-136.
(1993): Палеонтология и эволюция позвоночных. М.: Мир. 2: 1-280.
(1988): Расказы об экономии природы. Киев: Вища школа. 1-173.
(1985): Летающие ночью. М.: Знание. 1-159.
. (1979): Зоология позвоночных. М.: Высшая школа. 2: 1-271.
(1985): Демография птиц. Л.: Наука. 1-285.
(1957): Опыт эколого-систематического исследования эволюции птиц. - Вопр. экологии. Киев.: КГУ. 269-276.
(1955): К экологии серой куропатки в степных районах европейской части СССР. - Уч. зап. каф. зоол. Моск. гос. пед. ин-та. М. 38: 157-215.
(1974): Основы палеонтологии. М.: Мир. 1-390.
(1981): Введение в теорию эволюции. М.: МГУ. 1-318.
(1973): Систематика млекопитающих. М.: Высшая школа. 1-432.
(1980): Современные данные о происхождении птиц. - Орнитология. М.: МГУ. 15: 165-178.
, , (1986): Разведение редких видов птиц. М.: Агропромиздат. 1-206.
(1947): Из жизни птиц. М.: ИЛ. 1-214.
(1983): Скорость полета и размеры птиц. - Вестн. зоол. 6: 67-71.
Чельцов- (1982): Экология птиц. М.: МГУ. 1-127.
(1922): Курс сравнительной анатомии позвоночных животных. Москва-Петроград. 1-620.
(1938): Основы сравнительной анатомии. М.: Учпедгиз. 1-487.
Шмидт- (1978): Как работает организм животного? М.: Мир. 1-140.
Шмидт- (1987): Размеры животных: почему они так важны. М.: Мир. 1-259.
(1966): Морфоэкологические приспособления к скоростному полету у птиц. - Механизмы полета и ориентации птиц. М.: Наука. 64-81.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 |
