


На правах рукописи
Видообразование и систематика грызунов (Rodentia: Sciuridae, Cricetidae, Muridae)
по данным аллозимного анализа
03.00.08 – зоология
03.00.15 – генетика
Автореферат
диссертации на соискание ученой степени
доктора биологических наук
Владивосток - 2008
![]()
Работа выполнена в лаборатории эволюционной зоологии и генетики
Биолого-почвенного института Дальневосточного отделения Российской академии наук
Официальные оппоненты:
доктор биологических наук, старший научный сотрудник
доктор биологических наук, член-корреспондент РАН
доктор биологических наук, профессор
Ведущая организация:
Институт биологии развития
им. РАН (Москва)
Защита состоится 15 октября 2008 г. в 10 часов на заседании
диссертационного совета Д 005.003.03
при Биолого-почвенном институте ДВО РАН
Владивосток, проспект 100 лет Владивостоку, 159
С диссертацией можно ознакомиться в Центральной научной библиотеке ДВО РАН
Автореферат разослан «__» августа 2008 г.
Ученый секретарь диссертационного совета
кандидат биологических наук
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность темы исследования. Формализация в рамках иерархической системы классификации, осуществленная К. Линнеем (1758), создала основы для упорядочения биологического разнообразия. Это позволило Ж. Ламарку, Ч. Дарвину, Э. Геккелю и их последователям разработать классификацию биоты, основанную на филогенетических связях, где теория эволюции и систематика представляют собой взаимосвязанные стороны понимания происхождения, развития и современной организации окружающей нас живой природы. Понятие «вид», рассматриваемое первоначально только как «не изменяющаяся элементарная систематическая единица», оставаясь единицей систематики, приобрело смысл концептуальной единицы эволюции и базовой категории биоразнообразия определенного отрезка времени (Bock, 2004).
Майром (1968, 1971) биологическая концепция наиболее полно отражает обе стороны критерия вида (вид – филетическая линия от предков к потомкам, эволюционирующая отдельно от других таких же линий, и вид – представитель биоты конкретного периода времени). Направленная на выявление сущности вида (поддержание целостности хорошо сбалансированных гармоничных генных пулов) биологическая концепция, кроме того, определяет и пути поиска характеристик для выявления диагностических рамок (границ) конкретных видов – таксонов. Для бисексуальных диплоидных организмов, в том числе и рассматриваемых в работе представителей Rodentia, особое место занимают полиморфные кодоминантно наследуемые характеристики, позволяющие не только обнаружить различия таксонов, описать уникальность их геномов, но и исследовать общие эволюционные проблемы.
Именно такими признаками являются множественные молекулярные формы ферментов и других водорастворимых белков, которые активно исследуются после открытия метода зимограмм (сочетание гельэлектрофореза неочищенных тканевых гомогенатов с последующим гистохимическим окрашиванием на специфическую ферментативную активность) – первого метода генетического исследования на молекулярном уровне.
С первых работ (Harris, 1966; Hubby, Levontin, 1966) стало ясно, что анализ множественных молекулярных форм дополняет, расширяет и преобразует понимание микроэволюционных процессов, которое было достигнуто при исследовании надмолекулярных (морфологических, кариотипических и др.) характеристик и вносит существенный вклад в построение таксономической системы. Актуальным является всестороннее изучение уровня, характера белковой изменчивости и дифференциации у широкого спектра форм разной биогеографической принадлежности, экологии и эволюционной судьбы.
Реальная сложность процессов возникновения видов в природе предполагает многообразие моделей видообразования и картин дивергенции. Территория Евразии представляет собой уникальную «природную лабораторию» для исследования эволюционных преобразований ее биоты, а выбранные представители Sciuridae, Cricetidae и Muridae: суслики – Spermophilus, бурундук – Tamias, лесные полевки – Myodes, серые полевки – Microtus и домовые мыши – Mus поставляют обширный спектр форм для изучения внутривидовой географической изменчивости и генетической дифференциации в процессе своего эволюционного становления. В то же время более чем 200-летняя история их морфологического описания не привела к единому мнению по вопросам таксономии и систематики ряда форм ни на внутривидовом, ни на межвидовом уровнях. Поэтому актуальным является привлечение аллозимных маркеров для решения и этих задач.
Несмотря на стремительный прогресс молекулярной биологии, приведший к разработке широкого спектра методов анализа ядерной и митохондриальной ДНК, аллозимы продолжают оставаться востребованными генными маркерами. Причинами их использования являются огромная производительность метода зимограмм, позволяющая получить сведения о генотипическом составе популяций любых видов, причем каждый индивид легко может быть охарактеризован по нескольким десяткам генных локусов. Многообещающей для объяснения эволюции отдельных таксонов представляется возможность комбинированного использования метода высокого разрешения (секвенирование гена) и широкого охвата (географический анализ распространения электрофоретических фенотипов выявленных генных мутаций).
Цель и задачи исследования. Целью работы является изучение закономерностей генетической дивергенции (уровня и характера генетической дифференциации) для таксонов и филетических линий грызунов с различающимися спектрами таксономических и биогеографических характеристик.
Для достижения этой цели были поставлены следующие задачи.
1. На примере Spermophilus, Tamias (Sciuridae) исследовать внутривидовую генетическую дифференциацию широкоареальных материковых видов, межвидовые различия палеарктических таксонов.
2. На примере дальневосточных Cricetidae исследовать генетическую дифференциацию в условиях островной изоляции для фоновых и эндемичных видов (Myodes), а также в интродуцированных островных популяциях с известным возрастом изоляции (Microtus).
3. На примере дальневосточных Microtus – для которых можно предположить инициирующую роль хромосомных перестроек в видообразовании – рассмотреть особенности внутривидовой и межвидовой аллозимной дифференциации морфологически близких, но кариотипически различающихся таксонов.
4. Изучить аллозимную дифференциацию Mus musculus. На основе исследования белкового полиморфизма провести ревизию систематики фауны домовых мышей территории бывшего СССР. На основе литературных и собственных данных реконструировать схему заселения домовой мышью Евразии, в том числе территории бывшего СССР.
5. Для всех исследованных форм сформулировать таксономическую гипотезу с учетом полученных данных.
Научная новизна. В результате выполненных исследований изучена внутри - и межвидовая генетическая изменчивость 31 вида представителей 5 родов из 3 семейств грызунов. Для Spermophilus brevicauda, S. pallidicauda, разнохромосомных форм S. suslicus, Tamias sibiricus, Myodes rex, M. sicotanensis, Microtus maximowiczii, M. mujanensis, M. evoronensis, а также Microtus oeconomus Курильского архипелага и Mus musculus территории бывшего СССР это сделано впервые. Выделены три последовательных уровня генетической дифференциации, общих для представителей рассматриваемых семейств: уровень внутривидовой дифференциации, дифференциация эволюционно-молодых таксонов, как правило, дискуссионного таксономического ранга и дифференциация species bona. Тем самым показано, что видообразование шло по градуалистическому пути (кроме хромосомно полиморфных видов Microtus).
Теоретическое положение о возможности хромосомного видообразования (Воронцов, 1960; White, 1968) нашло реальное подтверждение в аллозимном исследовании хромосомно полиморфных дальневосточных видов рода Microtus maximowiczii, M. evoronensis, M. mujanensis.
Впервые для Rodentia, обитающих на территории бывшего СССР, проведено подробное исследование геногеографии белковых маркеров у видов с обширными материковыми и островными ареалами: Spermophilus undulatus, S. parryi, Tamias sibiricus, Myodes rutilus, M. rufocanus, Microtus oeconomus, M. maximowiczii, Mus musculus. Картины геногеографии отражают широкую представленность двух противоположных направлений эволюционного процесса: накопление различий между популяциями изолированных или пространственно разобщенных частей ареала и «размывание» дифференциации в результате вторичного контакта и последующей гибридизации (интрогрессии). Обнаружены и исследованы: в западном и южном Прибайкалье зона гибридизации западной и восточной групп подвидов длиннохвостого суслика, в северном Приамурье обнаружена зона гибридизации северной и южной групп популяций красной полевки, на территории юга Дальнего Востока России – зона гибридизации синантропных форм домовой мыши musculus и castaneus. Показано гибридное происхождение популяций домовой мыши Тувы, южных районов Читинская обл." href="/text/category/chitinskaya_obl_/" rel="bookmark">Читинской области.
Результаты исследования популяций небольших островов Курильской гряды предполагают значимость эффекта основателя, дрейфа генов и наличия свободных экологических ниш для начальных этапов дифференциации.
Обнаружено, что между обширными изолированными системами популяций формирование значимого уровня дифференциации может не происходить в течение длительного промежутка времени, превышающего десятки тысяч поколений (восточные изоляты длиннохвостого суслика; изоляты Myodes rutilus, M. rufocanus островов Сахалин" href="/text/category/sahalin/" rel="bookmark">Сахалин, Хоккайдо и прибрежных материковых территорий).
Показана возможность использования аллозимных характеристик для дифференциации видовых форм Spermophilus, Myodes, Microtus, Mus Палеарктики. Уточнены спорные вопросы их систематики. Подтверждены различия видового ранга у Spermophilus mayor, S. erythrogenys sensu stricto, S. pallidicauda, S. brevicauda; S. suslicus и S. odessanus; Myodes rufocanus, M. sikotanensis и M. rex. Обнаружены различия видового ранга у западной и восточной групп популяций Spermophilus undulatus. Показана необходимость повышения статуса Microtus maximowiczii gromovi c подвидового до вида Microtus gromovi.
На основании дифференциации по аллозимным характеристикам проведена ревизия систематики домовой мыши бывшего СССР. Обосновано существование на данной территории трех видов рода Mus: M. musculus, M. macedonicus и M. spicilegus. Пересмотрены их ареалы. Выдвинута и обоснована гипотеза о центре и направлениях расселения синантропных форм домовой мыши.
Положения, выносимые на защиту.
1. Внутривидовая и межвидовая структура аллозимной изменчивости и дифференциации адекватно отражает процесс и этапы эволюции таксона.
2. Аллозимные характеристики эффективны для решения таксономических задач на уровнях видов и подвидов Spermophilus, Tamias, Myodes, Microtus, Mus и выяснения истории формирования их ареалов.
3. Соотношение кариологической и аллозимной дифференциации может служить характеристикой эволюционной судьбы таксонов.
Теоретическое и практическое значение работы заключается в применении полученных данных в практике систематики и разработке актуальных проблем микроэволюции. Полученные данные использованы в ряде определителей и сводок по млекопитающим. Исследование геногеографии широкоареальных видов сусликов и аллозимное описание домовой мыши бывшего СССР оказались полезными для многих исследователей, о чем можно судить по довольно частому их цитированию. Внесенные уточнения в систематику грызунов следует учитывать в лекционных курсах по систематике млекопитающих, в чеклистах и определителях. Наши работы могут являться базой для сравнительных оценок при изучении этих групп Rodentia новыми молекулярно-генетическими методами
Таксономические исследования сусликов, полевок и домовых мышей практически значимы, т. к. эти грызуны являются компонентами многих биоценозов, в том числе и переносчиками особо опасных инфекций.
Апробация результатов. Результаты работы доложены на 1-м (Москва, 1974), 5-м (Рим, 1989), 9-м (Саппоро, Япония, 2005) Международных конгрессах по млекопитающим; на 14-м Международном генетическом конгрессе (Москва, 1978); на 2-м (Москва, 1978), 3-м (Москва, 1982), 4-м (Москва, 1985), 5-м (Москва, 1990), 6-м (Москва, 1999) съездах Всесоюзного териологического общества; на 5-м (Саратов, 1980), 7-м (Свердловск, 1988) Всесоюзных совещаниях по грызунам; на 5-м съезде ВОГИС (Москва, 1987); на 2-м симпозиуме по экологической генетике млекопитающих (Лодзь, Польша, 1994); на ежегодном совещании Японского общества маммологов (Саппоро, Япония, 1997); на 6-й Международной конференции «Грызуны и Пространство» (Хайфа, Израиль, 1998); на 1-м Международном симпозиуме по популяционной, эволюционной и экологической генетике животных (Владивосток, 1995); на Международном симпозиуме «Современные подходы в популяционной, эволюционной и экологической генетике» (Владивосток, 1998); на Международной конференции «Эволюция, генетика, экология и биоразнообразие» (Владивосток, 2000), на Международном симпозиуме «Вид и генетическое разнообразие диких животных восточной Азии» (Хайяма, Япония, 2000); на 4-ом (Брно, Чехословакия, 2003) и 5-м (Сиена, Италия, 2007) Европейских конгрессах по млекопитающим; на Сибирской зоологической конференции (Новосибирск, 2004), на Международной конференции «Современные проблемы биологической эволюции» (Москва, 2007).
Публикации. По теме диссертации опубликовано 79 работ..
Структура и объем диссертации. Диссертационная работа состоит из введения, 11 глав, заключения, выводов и списка цитированной литературы. Работа изложена на 301 странице, включает 59 таблиц и 37 рисунков. Список цитированной литературы состоит из 356 работ, в том числе 144 иностранных.
Благодарности. Мне хотелось бы выразить искреннюю признательность моему учителю Николаю Николаевичу Воронцову, оказавшему большое влияние на выбор области интересов и мою мировоззренческую позицию, глубокоуважаемым коллегам-соавторам, а также участникам многочисленных экспедиций, без которых это исследование было бы невозможным.
СОДЕРЖАНИЕ РАБОТЫ
Глава 1. МАТЕРИАЛЫ И МЕТОДЫ
В рамках диссертационной темы в аллозимное исследование были вовлечены представивида 5 родов, трех семейств (Sciuridae, Cricetidae, Muridae) грызунов Палеарктики. Для
2726 экз. грызунов из 273 географических точек экспериментальный анализ материала проведен автором. Конкретный объем этого материала (список видов и перечень выборок) представлен в 11 таблицах главы.
Для выявления генетической изменчивости использовался электрофорез белков в крахмальном геле или на целлюлозо-ацетатных пластинках с последующей окраской гелей и пластинок для выявления специфичной активности. Применялись буферные системы, условия электрофоретического разделения и методы окрашивания по Poulic, 1957; Kristiansson, 1963 в модификации ; Серов и др., 1977; Selander et al., 1971; Pasteur et al., 1988. В работе рассматривались до 26 интерпретационных локусов, кодирующих ферменты и неферментные белки следующего списка: лактатдегидрогеназа (LDH), малатдегидрогеназа (MOR), малик энзиим (MOD), α-глицерофосфатдегидрогеназа (GPD), 6-фосфоглюконатдегидрогеназа (PGD), глюкозо-6-фосфатдегидрогеназа (G6PD), изоцитратдегидрогеназа (IDH), сорбитолдегидрогеназа (SDH), ксантиндегидрогеназа (XDH), фосфоглюкомутаза (PGM), супероксиддисмутаза (SOD), аспартатаминотрансфераза (AAT), лейцинаминопептидаза (LAP), эстеразы (Es), гемоглобин и некоторые белки спектра общего белка плазмы крови.
Статистическая обработка экспериментальных данных осуществлялась стандартными методами вариационной математической статистики и специализированными методами, разработанными для анализа результатов электрофоретических исследований. При этом использовалось соответственно, стандартное и специализированное программное обеспечение. В главе приведено подробное описание используемых методов.
Построение дендрограмм проводилось на основе матрицы расстояний (генетических дистанций) методом попарного внутригруппового невзвешенного среднего (UPGMA) и на основе представленности аллельных вариантов методом ближайших соседей (NJ) с помощью специализированных пакетов прикладных программ BIOSYS-1 (Swofford and Selander, 1981) и PHYLIP 6,3 (Felsenstein, 1997).
Раздел 1. БЕЛКОВЫЙ ПОЛИМОРФИЗМ КАК ПОКАЗАТЕЛЬ ЭВОЛЮЦИОННЫХ ИЗМЕНЕНИЙ И ПОСТРОЕНИЙ СИСТЕМАТИКИ У НАЗЕМНЫХ БЕЛИЧЬИХ Marmotinae Pocock, 1923 (Rodentia, Sciuridae) ПАЛЕАРКТИКИ
Идеальным объектом для исследования эволюционных изменений, сопровождающих процесс видообразования, является род, в пределах которого существует достаточно большое число разнокачественных видов – от видов с незавершенной морфологической дивергенцией до несомненных species bona. Желательно, чтобы были представлены различные структуры видовых ареалов: а) по размерам – от локальных до обширных, б) по внутривидовой подразделенности – от сплошных до системы изолятов, разделенных непреодолимыми географическими барьерами, и
в) по взаимоположению ареалов различных видов – аллопатрические, парапатрические и т. д. Род Spermophilus Cuvier, 1825 соответствует всем вышеперечисленным требованиям и поэтому является ключевым объектом настоящего исследования.
Несмотря на то, что суслики относятся к наиболее интенсивно исследуемым группам животных и их морфологическая дифференциация находится под пристальным вниманием исследователей уже почти два века, сформировались противоречивые представления о числе и границах систематических единиц. Различными авторами в Палеарктике было выделено от 6 до 15 видов. Для внесения ясности в таксономию данной группы возникла необходимость привлечения иных, неморфологических дискриминирующих характеристик. Настоящий раздел посвящен описанию степени и характера белкового полиморфизма в палеарктических таксонах Spermophilus и сопоставлению уровней их генетической и морфологической дифференциации.
Глава 2. ГЕОГРАФИЧЕСКАЯ ИЗМЕНЧИВОСТЬ ДЛИННОХВОСТОГО СУСЛИКА (Spermophilus undulatиs Pallas, 1778) КАК РЕЗУЛЬТАТ ДИВЕРГЕНЦИИ В УСЛОВИЯХ ОБШИРНОГО ПАЛЕАРКТИЧЕСКОГО АРЕАЛА
Глава открывается подробным описанием ареала, основанным на кадастрово-справочной карте (Межова, Фрисман, 1984). Ареал характеризуется огромной протяженностью от Джунгарского Алатау на западе до Приамурья на востоке и от Якутии на севере до Монголии на юге, и значительной прерывистостью, т. е. наличием большого числа изолированных участков, из которых два крупных краевых - один на западе, а другой на севере.
Анализ литературных данных морфометрических и морфотипических признаков демонстрирует значимое отличие восточных подвидов S. u. menzbieri Ognev, 1937 и S. u. jacutensis Brandt, 1943 от западных S. u. stramineus Obolensky, 1927, S. u. eversmanni Brandt, 1841.
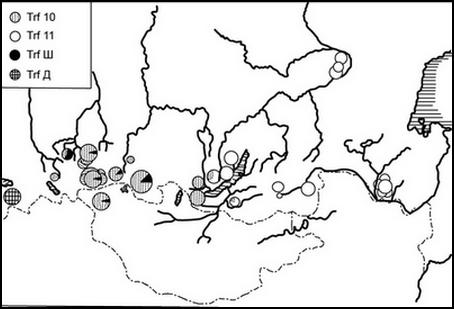 |
Географическая изменчивость встречаемости электрофоретических вариантов сывороточного белка трансферрина (Trf) показывает заселение огромных пространств популяциями со сходными (Западная–Центральная Сибирь) или идентичными (Забайкалье, Якутия, Приамурье) генотипами (Воронцов, Фрисман и др., 1978). В Прибайкалье нами обнаружена гибридная зона, где совместно встречаются аллели популяций различающихся регионов, в том числе и в гетерозиготном состоянии (рис. 1).
Рис. 1. Геногеография трансферринов длиннохвостого суслика
В главе рассматривается геногеография 14 локусов. Три из них − трансферрин, альбумин (Alb) и постальбумин (Postalb) − показывают полное различие аллельного состава в популяциях западного и восточного регионов и гибридный характер прибайкальских популяций (Фрисман, Воронцов, 1989). Анализ 25 локусов обнаружил значимый уровень дифференциации (см. табл. 1.) между «западной» и «восточной» формами.
Сопоставление геногеографической изменчивости и внутривидовой системы показывает, что подвидовой характер цветовых различий между S. u. stramineus и S. u. eversmanni, формирующих западную группу популяций, не подтверждается данными электрофореза белков, а морфологически различающиеся голоценовые изоляты восточной группы (S. u. intercedens Ognev, 1937, S. u. jacutensis и S. u. menzbieri), судя по результатам исследования 25 локусов, генетически близки. S. u. undulatus Pallas, 1773, объединяющий гибридную зону и соседние популяции, гетерогенен по признакам различных уровней. Отличительной чертой популяций гибридной зоны является неоднозначный результат определения их подвидовой принадлежности при привлечении нескольких характеристик: морфологических, морфотипических, иммунологических, генетических, хорошо «работающих» при разделении популяций западной и восточной частей ареала.
Заключительный параграф главы посвящен истории становления ареала вида. Данные палеонтологии показывают, что в Восточной Азии длиннохвостый суслик пережил эпохи нескольких оледенений (Ербаева, 1970; Вангенгейм, 1977). Палеогеографические, палеоэкологические (Adams, 1997) и генетические данные позволяют предположить длительный период разделения ареалов западной и восточной форм. Вырубка лесов и прокладка дорог, по-видимому, явилась тем антропогенным фактором, который привел к вторичному контакту в районах Предбайкалья и юго-западного Забайкалья и появлению гибридной зоны.
Глава 3. ГЕОГРАФИЧЕСКАЯ ИЗМЕНЧИВОСТЬ И ГЕНЕТИЧЕСКАЯ ДИФФЕРЕНЦИАЦИЯ БЕРИНГИЙСКОГО СУСЛИКА (Spermophilus parryi Richardson, 1925)
Формирование ареала S. parryi в большой степени связано с историей Берингии, что привлекает особое внимание к исследованию его внутривидовой изменчивости и дифференциации.
Изложение главы начинается с описания подвидового состава и ареала вида, который простирается от долины р. Яна в Евразии до Северо-Западной территории Канады.
Для исследования геногеографической изменчивости нами привлекались 14 интерпретационных локусов. Наиболее полные и интересные данные получены по сывороточному белку трансферрину, для которого к началу нашего исследования уже была известна геногеография в неарктической части ареала, что предоставляло возможность рассмотреть эволюционную историю вида в целом.
В палеарктической части ареала большинство рассматриваемых выборок оказались полиморфны по двум аллелям – Trf-6 и Trf-7. Лишь выборки восточной оконечности п-ова Чукотка оказались практически мономорфными по Trf-7 (рис. 2). В главе приводится изложение

Рис. 2. Геногеография трансферринов берингийского суслика
статистического исследования, показавшего плавный клинальный характер увеличения частоты аллеля Trf-7 от Верхоянья и п-ова Камчатка к п-ову Чукотка (Vorontsov, Frisman et al., 1980). Полученные данные по временным изменениям частот аллелей в некоторых популяциях не искажают картины клинальной изменчивости (Воронцов, Фрисман и др., 1983).
В популяциях неарктической части ареала Trf-6 и Trf-7 пространственно разобщены (Nadler, Hoffman, 1977). Генетическая дифференциация неарктических подвидов на арктическую и субарктическую формы сопровождается дифференциацией по морфологическим признакам, тогда как в Палеарктике представлена слабая клинальная изменчивость морфологических характеристик. Географическая изменчивость в популяциях неарктической части ареала берингийского суслика предполагает примат изоляционного и миграционного факторов в ее возникновении, тем самым подтверждая их значимость и в евразийских популяциях. Согласно палеонтологическим данным во время Сартанского оледенения берингийский суслик уже заселял Аляску (Repenning et al., 1964) и северо-восток Евразии (Виноградов, 1948). Благодаря существованию Канадского ледникового щита ареал берингийского суслика в Неарктике в то время представлял собой несколько изолированных участков, один из которых Берингийским мостом суши соединялся с евразийской частью ареала. Кажется возможным, что гомозиготизация по Trf-7 популяций п-ова Чукотка, о-ва Св. Лаврентия и юга п-ова Аляска имеет общие корни по своему происхождению в Берингийском рефугиуме. То есть 10 тыс. лет изоляции после возникновения Берингова пролива не привели к значимым различиям в морфологии (Чернявский, 1976) и не изменили их единообразия по рассматриваемому генетическому маркеру.
Западные (азиатские) границы Берингийского моста, в отличие от восточных (алясканских), в ледниковые периоды не были наглухо закрыты. Существовали межледниковые коридоры, соединяющие Берингийскую часть суши с остальной Азией. Однако эти коридоры в достаточно продолжительные периоды могли быть закрыты экологическими барьерами, такими как болотистые территории, препятствовавшие продвижению обитателей открытых пространств. Иными словами, в возникновении современной картины клинальной изменчивости встречаемости вариантов трансферрина в палеарктической части ареала значимое место мог занимать вторичный голоценовый контакт между изолятом Берингийского моста, гомозиготным по Trf-7, и основным массивом, по-видимому, изначально полиморфным по Trf-6 и Trf-7. Складывается впечатление, что первоначальное заселение палеарктической части ареала произошло полиморфными по Trf-6 и Trf-7 вселенцами.
Результаты генетического исследования длиннохвостого и арктического сусликов предполагают, что современные картины внутривидовой изменчивости формировались в течение не одного десятка тысяч лет. Определяющим фактором современных картин их внутривидовой дифференциации является динамика крупных изолирующих барьеров последнего ледникового цикла, приведшая в голоцене к объединению изолированных частей ареалов.
Глава 4. ВНУТРИВИДОВАЯ ГЕНЕТИЧЕСКАЯ ДИФФЕРЕНЦИАЦИЯ СУСЛИКОВ ЮГА ВОСТОЧНОЙ ЕВРОПЫ И ЮГА ЗАПАДНОЙ СИБИРИ
Ареалы сусликов этого региона парапатрические, либо разделены речными преградами. Существующие представления о числе и объеме таксонов зачастую противоречивы.
Крапчатый суслик Spermophilus suslicus Guldenstaedt, 1770 S. suslicus обитает на территории центральной и западной частей Европейской равнины. Внутривидовая морфологическая изменчивость этого вида проявляется в незначительных изменениях признаков в направлении север – юг (Громов, Ербаева, 1995). Вид представлен двумя хромосомными формами. Первая с 2n=36 распространена на запад от р. Днепр, а вторая с 2n=34 обитает на восток от Днепра до Волги (Воронцов, Ляпунова, 1970; Орлов и др., 1969; Кораблев, Ляпунова, 1996). Аллозимное исследование показало, что 10 из 25 интерпретационных локусов полиморфны. Для 6 из них не обнаружено ни значительной изменчивости, ни существенной дифференциации сравниваемых кариотипических форм. Два локуса показывают мозаичную картину распределения общих аллелей. Хромосомные формы различаются аллельным составом двух белков - Alb и Trf (Фрисман и др., 1999). Существующее подразделение крапчатого суслика на две кариотипические формы сопровождается дифференциацией этих форм на аллозимном уровне при большом сходстве популяций внутри каждой из форм. Такой результат предполагает постепенное накопление аллозимных различий между исследуемыми формами, что допускает достаточно длительный период раздельной эволюции их филетических линий. Современный временной срез этих линий наиболее полно соответствует понятию «потенциальные виды», предлагаемому Э. Майром (Mayr, 1996) для различающихся аллопатричных таксонов. По правилам приоритета, крапчатые суслики правобережья Днепра должны рассматриваться как S. odessanus Nordman, 1840, а левобережья – как S. suslicus sensu stricto Guld, 1770.
Малый S. pygmaeus Pallas, 1778 и кавказский горный S. musicus Menetries, 1832 суслики. Ареал малого суслика протяжен от среднего Приднепровья на западе до районов Целинограда, Караганды и западных окраин Муюнкума и Бетпак-Далы на востоке. Кавказский горный суслик - обитатель северных склонов центральных частей главного Кавказского хребта. Неопределенность таксономического ранга горного суслика обусловлена его слабыми морфологическими отличиями от малого суслика. различия Между S. рygmaeus и S. musicus обнаружены различия по характеристикам черепа (Голубев и др., 1976), строению отдельных хромосом (Воронцов и Ляпунова, 1969) и локализации гетерохроматина (Кораблев, 1983). Различия лево - и правобережных популяций малого суслика, при сходстве последних с S. musicus, показаны при исследовании контрольного фрагмента С региона мтДНК (Ермаков и др., 2006) и по признакам звукового сигнала извещающего об опасности (Никольский и др., 2007).
Аллозимное сравнение горного суслика с малым сусликом левобережья Волги показало дискриминирующее различие этих форм только по одному локусу (Trf) при близком их сходстве либо идентичности по 24-м локусам. Ранее по этому белку были обнаружены подобные различия между горным сусликом и малым сусликом правобережья (Nadler et al., 1974). Судя по шкале относительной подвижности вариантов трансферринов, разработанной Ч. Надлером с соавторами (1974) для Spermophilus, право - и левобережные выборки малого суслика имеют различающиеся варианты Trf.
Существующий объем данных позволяет предположить самостоятельные эволюционные линии для трех изолятов: горный суслик и два изолята малого суслика, разделенные р. Волга.
Определенность оценки таксономического ранга этих форм может быть достигнута лишь при выяснении уровня их репродуктивной изоляции. Таксономическая интерпретация еще более осложнена тем фактом, что малый суслик способен вступать в гибридизацию даже с представителями, несомненно, самостоятельных видов. Так, на правом берегу Волги известны гибриды с крапчатым сусликом (Денисов, 1961), а на левобережье – с большим сусликом (Бажанов, 1944; Формозов, Никольский, 1986).
Суслики группы «major». Группа «major» объединяет большого, краснощекого и бледнохвостого сусликов, которые заселяют степные и полупустынные биотопы от Волги до Внутренней Монголии и от Новосибирская обл." href="/text/category/novosibirskaya_obl_/" rel="bookmark">Новосибирской области до Синьцзяна. На огромных пространствах внутри указанной территории суслики отсутствуют. Фактически ареал представляет собой серию больших и малых изолятов, разделенных веером крупных географических и экологических барьеров. В настоящее время не существует единого мнения ни о числе, ни об объеме видов в этой группе. На основании анализа морфологических характеристик выделяют либо один вид S. major Pallas, 1779 (Бобринский и др., 1965; Кузнецов, 1975), либо два вида: S. major, включающий erythrogenys Brandt, 1841 в ранге подвида, и S. pallidicauda Satunin, 1903 (Ognev, 1947; Ellermann, Morrison-Scott, 1951) или S. major и S. erythrogenys, включающий pallidicauda как подвид (Громов и др., 1965), либо три вида: S. major, S. erythrogenys и S. brevicauda, Brandt, 1943 (=intermedius Brandt, 1944) с подвидом S. b. pallidicauda (Огнев,1947; Слудский и др., 1969). На основе различий в хромосомных наборах последний был выделен в самостоятельный вид S. pallidicauda (Орлов и др., 1978).
Исследование 25 интерпретационных локусов проводилось нами на примере трех групп выборок из северо-западной (Ульяновская и Саратовская обл." href="/text/category/saratovskaya_obl_/" rel="bookmark">Саратовская области), южной (Казахстан) и юго-восточной (Монголия) частей ареала группы «major» (Кораблев, Фрисман и др., 2003). Кроме того, рассматривались выборки северо-восточной (Новосибирская обл.) и центральной (Павлодарская обл." href="/text/category/pavlodarskaya_obl_/" rel="bookmark">Павлодарская обл.) частей ареала. Четыре из десяти полиморфных локусов имели общие аллели во всех рассматриваемых таксонах. Как правило, этот аллель был основным или единственным в каждой из выборок. По Hb, G6pd, Alb, Trf, эстеразе плазмы-1 (Es-1), эстеразе гемолизата-1 (Es-g-1) географически близкие выборки имели сходное распределение аллелей. Между географически разобщенными группами выборок обнаружено отсутствие общих аллелей, что не позволяет объединять представителей группы «major» в составе единого вида S. major.
Много разногласий вызывал вопрос о расположении границы между большим и краснощеким сусликом. Наиболее спорна видовая принадлежность сусликов Тоболо-Иртышского междуречья. Результаты нашего биохимического исследования позволяют утверждать, что Тоболо-Иртышское междуречье заселяют суслики, близкие к южно-казахстанской форме, т. е. граница проходит по р. Тобол.
Большинство систематиков рассматривают большого и краснощекого сусликов в качестве самостоятельных видов, несмотря на широкую гибридную зону на стыке их ареалов. Аллозимное сравнение S. major с S. erythrogenys sensu lato показало фиксацию различий по Es-1, высокие частотные различия по Trf. Тем самым биохимические данные указывают на самостоятельность филетической линии S. major по отношению к S. erythrogenys sensu lato. Более значим уровень дифференциации, обнаруживаемый при сравнении S. major с выборками S. erythrogenys sensu lato отдельных регионов: северо-востока (S. e. erythrogenys), юга (S. e. pallidicauda), юго-запада (S. e. brevicauda). Фиксация различий касается 2-5 локусов, а состав таких локусов меняется от таксона к таксону.
Аллозимный анализ показывает отсутствие общих аллелей для 3 локусов у сусликов из северных и южных территорий, тем самым подтверждая правомочность представлений и о выделении S. brevicauda в качестве самостоятельного вида. Видовой статус короткохвостого суслика (brevicauda) подтверждается и данными исследования цитохрома b мтДНК (Ермаков и др., 2002).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
