


Для сусликов Монголии и Казахстана не обнаружено фиксированных различий ни по одному из исследованных локусов. Однако для двух локусов (Es-g-2 и Trf) сходство ограничивается лишь общими редкими аллелями.
Таксоны группы «major» имеют уровень аллозимной дифференциации, равный или превышающий дифференциацию между разнохромосомными формами крапчатого суслика, между горным и малым сусликом и между «западной» и «восточной» формами длиннохвостого суслика. Генетические (аллозимные и кариологические) различия подтверждают правомочность наиболее дробного подразделения, а именно, выделение в качестве единиц современного биоразнообразия самостоятельных таксонов S. mayor, S. erythrogenys, S. brevicauda и S. pallidicauda. В то же время очевидно, что процесс видообразования в данной группе не завершен. Дифференциация между геномами по морфологическим и генетическим характеристикам произошла, но процесс формирования репродуктивной изоляции незакончен. Подтверждение тому - существование широкой гибридной зоны между большим и краснощеким сусликами. Однако повышенная эмбриональная смертность и существенная диспропорция в половой структуре гибридов (Стариков, Жилин, 1999), несомненно, указывает на присутствие элементов их репродуктивной (постзиготической) изоляции.
Глава 5. АЛЛОЗИМНАЯ ДИФФЕРЕНЦИАЦИЯ, ГЕНЕТИЧЕСКИЕ СВЯЗИ И СИСТЕМАТИКА СУСЛИКОВ РОДА Spermophilus ПАЛЕАРКТИКИ
В данной главе основное внимание уделено описанию характера межвидовой аллозимной дифференциации палеарктических Spermophilus, рассматриваются спорные вопросы систематики и эволюционная история рода.
Межвидовая дифференциация. В исследование межвидовой аллозимной дифференциации, кроме рассматриваемых выше видов, были включены S. fulvus Lichtenstein, 1825, S. citellus Linnaes,1766, S. xanthoprimnus Benneth, 1825, S. relictus Kashkarov, 1823 и S. dauricus Brandt, 1844, т. е. практически все (за исключением S. alaschanicus Buchner, 1888) палеарктическиe виды рода Spermophilus. Большая часть исследованных локусов (16 из 26), показала фиксацию альтернативных аллелей в выборках, принадлежащих к разным видам. Число таких локусов между сравниваемыми парами видов изменяется от 1 до 13. Генетические дистанции (по Nei, 1978) таксонов видового ранга изменяются от 0,06, что соответствует уровню подвидовых различий, до 0,712 − величина, приближающаяся к максимальным значениям этого показателя, известного для грызунов (Фрисман, Павленко, 1987). Парапатрические и викариирующие виды обладают меньшими значениями дистанций, чем аллопатрические, пространственно разобщенные. В главе проводится сравнение генетической дифференциации на популяционном, подвидовом и видовом уровнях. Особое внимание уделяется формам спорного таксономического ранга.
Табл. 1. Генетическая дивергенция Spermophilus Палеарктики
Группы сравнения | Число пар сравнений | Размах значений генетических дистанций D (Nei, 1978); среднее значение (довер. интервал) | Число дискри-минирующих локусов (из 26) | |
1 | Популяции | 21 | 0-0,053; 0,017 (0,006) | 0 |
2 | Подвиды | 13 | 0,004-0,085; 0,037 (0,011) | 0 |
3 | species bona (=«хорошие» виды) | 139 | 0,162- 0,712; 0,386 (0,023) | 2-13 |
Спорные таксоны (виды – группы подвидов?) | ||||
4 | S. odessanus - S. suslicus | 16 | 0,081- 0,165; 0,115 (0,011) | 2 |
5 | Таксоны группы «major» | 56 | 0,064-0,276; 0,164 (0,016) | 0-4 |
6 | S. musicus - S. pygmaeus | 2 | 0,06 | 1 |
7 | S. undulatus «запад» – «восток» | 2 | 0,191-0,197; 0,194 | 2 |
8 | S. undulatus - S. parryi | 2 | 0,355-0,360 | 8 |
9 | S. citellus –S. xanthoprymnus - S. relictus - S. dauricus | 9 | 0,24 –0,614; 0,423 (0,069) | 5-7 |
Анализ аллозимной дифференциации обнаруживает у палеарктических Spermophilus разные этапы процесса становления и развития видов (Frisman, Korablev, 2000). Наименьший уровень различий характеризует популяции и географически близкие подвиды (строки 1-2 табл. 1). Пары генетически близких видов: S. suslicus и S. odessanus, S. pygmaeus и S. musicus, - с дискриминирующими аллозимными характеристиками, несомненно, представляют собой независимые филетические линии, однако, в силу аллопатричности ареалов, их таксономический ранг в настоящее время не определяется однозначно. Майру (1996) мы рассматриваем их как «потенциальные» виды. Таксоны надвидовой группы «major»и географические формы
S. undulatus представляют собой иной пример. Настоящее время – это период проверки «защищенности геномов» членов групп генетически, и не только генетически, различающихся и ранее независимо эволюционировавших. Уровень аллозимных различий S. citellus, S. хanthoprymnus, S. relictus, S. dauricus, объединяемых некоторыми систематиками в единый вид S. citellus, сопоставим с таковым в группе «хорошие» виды, что несомненно указывает на самостоятельный видовой ранг рассматриваемых таксономических форм. Арктический и длиннохвостый суслики – «виды с незавершенной дифференциацией морфологических различий» – также характеризуются высоким уровнем аллозимной дифференциации.
Эволюционная история рода по результатам генетических исследований. Современный сценарий палеонтологических реконструкций происхождения мировой фауны сусликов предполагает, что суслики колонизировали Евразию в миоцене (Black, 1972; Qui, 1991). Суслики трибы Citellini на Евразийском континенте известны с фаун позднего виллафранка (2,5 млн. лет) (Агаджанян, 2005).
Накопленные к настоящему времени генетические данные, дополняя друг друга, создают достаточно стройную схему эволюции Spermophilus. Большой размах значений межвидовых генетических дистанций предполагает неодинаковый эволюционный возраст исследуемых таксонов, т. е. несколько вспышек видообразования. Наиболее близкая к нашему времени − разделение эволюционных линий pygmaeus - musicus; odessanus - suslicus; brevicauda - pallidicauda. К этому же периоду относится дивергенция географических форм длиннохвостого суслика.
Схема кариологической дивергенции рода, предложенная и (1970), касается более раннего, по-видимому эоплейстоценового, этапа эволюции рода. Эта схема предполагает одну общую эволюционную линию для современных 36-хромосомных видов: S. pygmaeus - S. musicus - S. major - S. erythrogenys - S. fulus - S. relictus - S. dauricus. Биохимические данные включают в эту эволюционную линию S. suslicus. Именно к этой эволюционной линии относится и «недавно» отделившиеся 34-хромосомные S. odessanus и S. pallidicauda. Аллозимное отличие S. relictus relictus может определяться структурой его ареала, обуславливающей более быструю скорость эволюции в «заоблачных островных» системах, и не противоречит включению этого таксона в филетическую линию 36 хромосомных видов. Самостоятельность линии citellus – хanthoprymnus подтверждается как кариологическими так и аллозимными данными.  Согласно кариологической схеме, S. undulatus и S. parryi формируют независимые линии, включенные в эволюционные группы северо-американских таксонов. Результаты исследования одного из локусов мт генома - полной последовательности цитохрома b (Harrison et al., 2003) согласуются с таким выводом. Подтверждая независимость эволюционных линий S. undulatus и S. parryi биохимические данные неожиданно объединяют последнего в единую эволюционную линию с S. dauricus. Несколько обособленное положение S. dauricus по отношению к 36-хромосомным палеарктическим видам показано и в выше обозначенном исследовании мт ДНК.
Согласно кариологической схеме, S. undulatus и S. parryi формируют независимые линии, включенные в эволюционные группы северо-американских таксонов. Результаты исследования одного из локусов мт генома - полной последовательности цитохрома b (Harrison et al., 2003) согласуются с таким выводом. Подтверждая независимость эволюционных линий S. undulatus и S. parryi биохимические данные неожиданно объединяют последнего в единую эволюционную линию с S. dauricus. Несколько обособленное положение S. dauricus по отношению к 36-хромосомным палеарктическим видам показано и в выше обозначенном исследовании мт ДНК.
Глава 6. ГЕОГРАФИЧЕСКАЯ ИЗМЕНЧИВОСТЬ АЗИАТСКОГО БУРУНДУКА Tamias sibiricus Laxmann., 1769 (RODENTIA, SCIURIDAE)
Tamias sibiricus населяет территорию от северо-востока европейской части России до Тихоокеанского побережья Азии. Представлен изолятами на островах Сахалин" href="/text/category/sahalin/" rel="bookmark">Сахалин, Кунашир и Хоккайдо. Все исследователи отмечают спорность как объема, так и состава 5-6 описываемых подвидов бурундуков, которые различаются друг от друга лишь по варьирующим цветовым признакам (Огнев, 1940; Формозов, 1928; Зубчанинова, 1962; Телегин, 1969). Оценка генетического сходства по 17 локусам показывает генетическую близость 6 выборок пространственно разобщенных локалитетов, подтверждая крайне слабое отличие подвидов сибирского бурундука друг от друга (Фрисман и др., 1984). По каждому из 3 полиморфных локусов один и тот же основной аллель встречается во всех выборках. Уровень генетической дивергенции подвидов сибирского бурундука сопоставим с таковым восточных подвидов длиннохвостого суслика и географически близких популяций остальных исследованных видов сусликов. Эти данные предполагают, что заселение бурундуком обширного ареала вслед за продвижением лесов на север после последнего оледенения происходило из одного рефугиума. При этом времени, прошедшего после данного события, не хватило для возникновения сколько-либо значимой генетической дифференциации подвидов этого высокоподвижного и обладающего относительно стабильной динамикой численности вида. Как не хватило времени и для возникновения генетической дифференциации между восточными подвидами длиннохвостого суслика, разделенными лесными массивами голоценового периода.
Исследование белкового полиморфизма наземных беличьих позволило рассмотреть формирование геногеографической изменчивости на обширных просторах северной и умеренной зон Евразии у Spermophilus, обитателей открытых пространств, и Tamias sibiricus, населяющих зону лесов. В то же время среди рассматриваемых наземных беличьих в островные системы распространяются лишь ареалы берингийского суслика и сибирского бурундука. Островная изоляция в несколько тысяч лет не оказала значимого влияния на внутривидовую дифференциацию этих видов.
Раздел 2. ГЕНОГЕОГРАФИЧЕСКАЯ ИЗМЕНЧИВОСТЬ И ГЕНЕТИЧЕСКАЯ ДИФФЕРЕНЦИАЦИЯ В УСЛОВИЯХ ОСТРОВНОЙ ИЗОЛЯЦИИ НА ПРИМЕРЕ ПОЛЕВОК РОДОВ Microtus SHRANK, 1738, Myodes PALLAS, 1811 (CRICETIDAE) ДАЛЬНЕГО ВОСТОКА РОССИИ
В геологическом прошлом колебания уровня океана, обусловленные неотектоническими процессами и крупными оледенениями приводили к значимым изменениям очертаний восточной окраины евразийского материка. Во времена регрессий уровня океана некоторые из ныне существующих островов входили в состав материка либо соединялись мостами суши между собой. Все эти изменения не могли не отразиться на генетической структуре видов, заселяющих данную территорию. Известный возраст островных изолятов позволяет рассматривать зависимость генетической дифференциации от времени изоляции. Для описания формирования генетической дифференциации в процессе островной изоляции привлечен Cricetidae, имеющий наиболее массовое представительство (как по масштабам распространения, так и по таксономическому составу) в дальневосточной островной фауне грызунов. Анализ изменчивости и генетической дифференциации 5 таксонов: Microtus oeconomus Pallas, 1776, Myodes rutilus Pallas, 1779, M. rufocanus Sundevall, 1846, Myodes sikotanensis Tokuda, 1935, M. rex Imaizumi, 1971 − проводится по единой схеме. Рассматривается геногеографическая изменчивость. Определяется степень накопления генетических различий в ряду: а) популяции различных частей одного и того же острова б) популяции, разделенные длительно существующими морскими преградами, в) подвидовые формы, представленные выборками из популяций, разделенных значительным расстоянием.
Глава 7. ГЕНОГЕОГРАФИЧЕСКАЯ ИЗМЕНЧИВОСТЬ И ГЕНЕТИЧЕСКАЯ ДИФФЕРЕНЦИАЦИЯ ПОЛЕВКИ-ЭКОНОМКИ Microtus oeconomus Pallas, 1776 КУРИЛЬСКИХ ОСТРОВОВ
На Дальнем Востоке Евразии полевка-экономка распространена от побережий Северного Ледовитого океана до северных районов Хабаровский край" href="/text/category/habarovskij_kraj/" rel="bookmark">Хабаровского края, обитает на п-ове Камчатка, Северных Курилах. Широко распространена она и по Центральным Курильским островам. Согласно запискам купца Г. Шелихова (Шелихов, 1971) и сведениям (Клумов, 1960), полевка-экономка появились на Центральных Курилах в конце девятнадцатого века. Исходным материалом для заселения послужили популяции Cеверных Курильских островов: Парамушира и Шумшу. Следовательно, возраст изоляции популяций полевки-экономки Курильских островов лишь слегка превышает 100 лет. Северные Курильские острова и Камчатка были разделены проливами около 10 тысяч лет тому назад (Велижанин, 1976; Безверхий и др., 2002). Таким образом, полевка-экономка предоставляет уникальную возможность наблюдения за самыми ранними этапами островной дифференциации природных популяций, а также возможность оценки уровня генетической дифференциации популяций при известном времени их изоляции. Для характеристики генетической дифференциации использовалось сравнение изменчивости по 24 интерпретационным локусам популяций 10 островов Курильской гряды (Шумшу, Парамушир, Атласова, Онекотан, Харимкотан, Шиашкотан, Матуа, Расшуа, Кетой, Симушир), полуострова Камчатка и Магаданская обл." href="/text/category/magadanskaya_obl_/" rel="bookmark">Магаданской обл. В главе приводится подробное описание географической изменчивости каждого из обнаруженных полиморфных локусов. Типичной картиной геногеографической изменчивости Pgd, Ldh-2, Idh-1, Pgm, Mod-1, Sod-1, Sod-2, Es-3 является мономорфность большинства выборок (островных и материковых) по одному и тому же аллелю, который в то же время является основным и в полиморфных выборках. Дополнительные варианты распределены мозаично. Лишь один локус (Es-1) и только в двух популяциях (окр. г. Магадан и о-в Онекотан) показывает смену основного аллеля. Суммарный набор уникальных аллелей в интродуцированных популяциях Центральных Курильских островов оказался выше, чем в популяциях Магаданской обл., п-ова Камчатка и Северных Курил. Присутствие уникальных аллелей в популяциях Центральных Курил, по-видимому, является первым шагом дивергенции островных популяций. При формировании популяций Центральных Курильских островов несомненно большое значение имели эффект основателя и дрейф генов. Тем не менее можно предположить, что увеличению аллельного разнообразия способствовало заселение новых территорий и освоение большого числа свободных ниш, что происходило на Центральных Курильских островах (Фрисман и др., 2003).
Cреднее значение величин дистанций D (Nei, 1978), наблюдаемых между популяциями одного и того же острова, статистически достоверно ниже наблюдаемых при межостровных сравнениях. Однако статистическая достоверность не подтверждается при использовании хордовых дистанций и дистанций Превости. Этот же тест указывает на высокую вероятность равенства средних дистанций Превости и хордовых, обнаруживаемых при сравнении изолятов п-ова Камчатка и островов Курильского архипелага. Эти данные являются подтверждением принадлежности популяций полевки-экономки п-ова Камчатка и Курильских островов к единому подвиду M. о. kamtschaticus Pallas, 1779. Сравнение межподвидового (M. o. koreni Allen, 1914 и M. o. kamtschaticus) и внутриподвидового (M. o. kamtschaticus) среднего значения генетических дистанций показало статистически достоверное (с вероятностью более 0,99) превышение величины первого над вторым. Данный вывод получен при анализе трех рассматриваемых дистанций.
Полученные оценки аллозимной дифференциации полевки-экономки п-ова Камчатка и Курильских островов, наряду с известными оценками возраста изоляции островов, предоставили возможность определить возраст расхождения камчатской и магаданской популяций как 0,14x106 лет, что не противоречит представлениям отечественных териологов о заселении п-ова Камчатка полевкой-экономкой в позднем плейстоцене (Костенко, 2000).
Глава 8. ВЛИЯНИЕ ОСТРОВНОЙ ИЗОЛЯЦИИ И ПРОСТРАНСТВЕННОЙ РАЗОБЩЕННОСТИ НА АЛЛОЗИМНУЮ ДИФФЕРЕНЦИАЦИЮ ФОНОВЫХ И ЭНДЕМИЧНЫХ ВИДОВ ЛЕСНЫХ ПОЛЕВОК Myodes Pallas 1811(=Clethrionomys Tilesius, 1850) ДАЛЬНЕГО ВОСТОКА РОССИИ
Myodes rutilus Pallas, 1779. У красных полевок с территории, охватывающей п-ов Камчатка, юг Магаданской обл. (M. r. jochelsoni J. Allen, 1903), острова Хоккайдо, Сахалин (M. r. mikado Thomas, 1905) и побережье Татарского пролива (M. amurensis Schrenck, 1858), по большинству полиморфных локусов (Ldh-1, Ldh-2, Mor-2, Mod-1, Mod-2, Mod-3, Xdh, Pgd, Sdh, Gpd, Lap и Trf) во всех выборках присутствовал один и тот же основной (с р>0,5) аллель, а географическая изменчивость проявлялась в локализации дополнительных аллелей (Фрисман и др., 2002). Наиболее значимая геногеографическая изменчивость – смена основных аллелей между «северной» (Магадан и Камчатка) и «южной» (острова и побережье Татарского пролива) группами популяций – была обнаружена по гемоглобину и G6рd.
Суммирование собственных и литературных данных геногеографии гемоглобина показывает, что в «северную» группу входят популяции Аляски, Северо-Востока России, Якутии и Среднего Амура. В «южную» группу объединяются популяции Амурского лимана, материкового побережья Татарского пролива, восточных отрогов Сихотэ-Алиня и островов Сахалин, Хоккайдо, Большой Шантар ((Nadler et al., 1978; Коробицына и др., 1983; Курышев, 1990; Фрисман и др., 2001). Можно предположить, что существующая картина сложилась, по меньшей мере, до отделения о-ва Сахалин от материка, т. е. более 7 тыс. лет тому назад. Древним географическим барьером между «северной» и «южной» формами, по-видимому, являлась долина пра-Амура, включающая в себя территорию современной Эворон-Чукчагирской впадины. При изменении русла появилась возможность соединения ареалов «северной» и «южной» форм и как результат, возможность появления гибридных популяций. Такая популяция обнаружена нами в окрестностях оз. Эворон.
Несмотря на достаточно пеструю картину распределения дополнительных аллелей, присутствие одного и того же аллеля в качестве основного или единственного определяет невысокий уровень дифференциации популяций внутри «южной» и «северной» групп при более высоком уровне дифференциации между группами (D (Nei, 1978) от 0,007 до 0,026 и 0,115 соответственно). Аллозимные данные подтверждают выделение М. r. jochelsoni и родство красных полевок островов Хоккайдо, Сахалин, Б. Шантар, Амурского лимана и восточных склонов Сихотэ-Алиня тем самым показывая комплексный характер М. r. amurensis. В состав этого подвида, кроме представителей «южной» гемоглобиновой группы (обитатели о-ва Большой Шантар, Амурского лимана и востока Приморья), включены популяции, принадлежащие к «северной» гемоглобиновой форме. Комплексный состав М. r. amurensis обнаружен (2000) на основании различий по морфологическим показателям тела и черепа. Иная картина изменчивости обнаружена М. Ивасой с соавторами (Iwasa et al., 2002) при анализе мт генома. Они объединяют в единую группу полевок п-ова Камчатка (M. r. jochelsoni) и о-ва Сахалин (M. r. amurensis) и выделяют в самостоятельную филетическую линию полевок о-ва Хоккайдо (M. r. micado).
M. rufocanus Sundervall, 1846. У красно-серой полевки островов Сахалин, Итуруп, Полонского, Хоккайдо (С. ruf. smithi Tomas, 1905), п-ова Камчатка и окрестностей г. Магадан (M. ruf. wosnessenskii Poliakov, 1881), так же как и у красной полевки, по большинству полиморфных локусов (Mor-2, Mod-1, Mod-2, Pgd, Gpd, Lap, Alb и Trf) во всех выборках присутствовал один и тот же основной аллель, а географическая изменчивость проявлялась в мозаичной локализации дополнительных аллелей. Два локуса показывают более высокий уровень географической изменчивости. По Es-4 обнаружена фиксация различных аллелей в островных популяциях. Mod-3 показывает отличие популяций Магадана и Камчатки от всех остальных. Значения несмещенных генетических дистанций 0,007-0,115 оказались «рамочными» и для внутривидовой генетической дифференциации M. rufocanus. Наиболее значимые различия обнаруживается между островными и пространственно удаленными от них (Магадан, Камчатка) популяциями.
Результаты аллозимного анализа согласуются с подвидовым разделением красно-серой полевки (по: Каталог…, 1981; Громов, Ербаева, 1995) в части объединения в единый подвид популяций северо-востока Азии, включая Камчатку, и выделения единого подвида для островных изолятов. Согласуется такое разделение и с результатами кариологического анализа (Воронцов и др., 1978). Еще одно подтверждение такому подразделению получено М. Ивасой и др. (Iwasa et al., 2000) при исследовании Sry гена, расположенного на Y хромосоме. По результатам секвенирования авторы сгруппировали дальневосточных красно-серых полевок в две формы – континентальную и островную (Сахалин – Хоккайдо).
Близость уровней внутривидовой генетической дифференциации красной и красно-серой полевок в рассматриваемых островных системах предполагает одновременность (в эволюционном смысле) заселения этими видами данного региона. Изолированное существование лесных полевок на островах в течение 7 тыс. лет (отделение Сахалина от материка) и 10 тыс. лет (Сахалина от Хоккайдо) не привело к статистически достоверному превышению величины аллозимной дифференциации популяций разных островов по сравнению с дифференциацией популяций различных частей о-ва Сахалин.
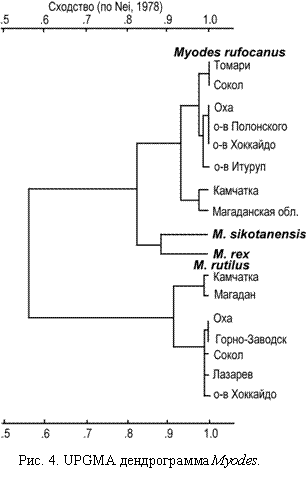 M. sikotanensis Tokuda, 1935 и M. rex Imaizumi, 1971. Шикотанская полевка, описана М. Токудой (1941) с о-ва Шикотан позднее сведена им же в синоним красно-серой полевки M. rufocanus bedfordiae, Tomas, 1905, а затем восстановлена с первоначальным названием M. sikotanensis (Бобринский и др., 1965). и (1978, 1990) отнесли сахалинскую форму к номинальному подвиду M. s. sikotanensis Tokuda, 1935. Запутанность таксономических взаимоотношений sikotanensis - rufocanus усугубляется присутствием описанного с о-ва Риcири и обитающего на о-ве Хоккайдо M. rex. Imaizumi, 1971. Ота (Ota, 1954) прямо придерживается взглядов М. Токуды (Tokuda, 1941) и сводит M. sikotanensis в синоним к M. ruf. bedfordiae, то Абе (Abe, 1968, 1973, 1973a) говорит о двух формах красно-серой полевки: Д-форме M. sikotanensis и М-форме M. rufocanus, - указывая, что они образуют на о-ве Хоккайдо по морфологии и экологии отчетливые локальные вариации, и считает, что шикотанская полевка должна быть сведена в синоним к M. rex (Study …, 1984; Abe, 2000). M. rex представлен как самостоятельный вид в сводке Г. Корбета (Corbet, 1978), однако другие авторы (Aimi, 1980; Павлинов, Россолимо, 1987) рассматривают его в составе M. rufocanus. Филогенетические исследования вариаций ядерной (рибосомальной) и митохондриальной ДНК показали, что M. rex и M. rufocanus представляют собой независимые филетические линии, разошедшиеся в раннем плейстоцене (Wakana et al., 1996), а наблюдаемые различия позволяют рассматривать их в качестве сестринских видов.
M. sikotanensis Tokuda, 1935 и M. rex Imaizumi, 1971. Шикотанская полевка, описана М. Токудой (1941) с о-ва Шикотан позднее сведена им же в синоним красно-серой полевки M. rufocanus bedfordiae, Tomas, 1905, а затем восстановлена с первоначальным названием M. sikotanensis (Бобринский и др., 1965). и (1978, 1990) отнесли сахалинскую форму к номинальному подвиду M. s. sikotanensis Tokuda, 1935. Запутанность таксономических взаимоотношений sikotanensis - rufocanus усугубляется присутствием описанного с о-ва Риcири и обитающего на о-ве Хоккайдо M. rex. Imaizumi, 1971. Ота (Ota, 1954) прямо придерживается взглядов М. Токуды (Tokuda, 1941) и сводит M. sikotanensis в синоним к M. ruf. bedfordiae, то Абе (Abe, 1968, 1973, 1973a) говорит о двух формах красно-серой полевки: Д-форме M. sikotanensis и М-форме M. rufocanus, - указывая, что они образуют на о-ве Хоккайдо по морфологии и экологии отчетливые локальные вариации, и считает, что шикотанская полевка должна быть сведена в синоним к M. rex (Study …, 1984; Abe, 2000). M. rex представлен как самостоятельный вид в сводке Г. Корбета (Corbet, 1978), однако другие авторы (Aimi, 1980; Павлинов, Россолимо, 1987) рассматривают его в составе M. rufocanus. Филогенетические исследования вариаций ядерной (рибосомальной) и митохондриальной ДНК показали, что M. rex и M. rufocanus представляют собой независимые филетические линии, разошедшиеся в раннем плейстоцене (Wakana et al., 1996), а наблюдаемые различия позволяют рассматривать их в качестве сестринских видов.
Наши исследования (Фрисман и др., 2002) показали, что как M. rex, так и M. sikotanensis имеют фиксированные различия по двум локусам (Xdh, и Trf) с M. rufocanus и по 6 локусам (Trf, Idh (-1, -2), Aat-1, Mod-2, Es-1) с M. rutilus, т. е. ведут себя как единое целое по отношению к рассматриваемым видам. В то же время, эндемичные таксоны различались между собой намного более чем островные популяции каждого из этих фоновых видов. Эндемики не имеют ни одного общего аллеля лейцинаминопептидазы. Кроме того, из обнаруженных у них 5 полиморфных локусов у двух локусов основной аллель полиморфного таксона не выявлен у другого таксона. Для Mod-3 в выборках представлены различные основные аллели. Аллозимная UPGMA дендрограмма (рис. 4) отражает подразделенность «северной» и «южной» региональных групп популяций красной полевки и иллюстрирует видовой ранг различий M. rufocanus, M. sikotanensis и M. rex.
M. rutilus и M. rufocanus являются фоновыми экологически пластичными видами. На рассматриваемой части ареала они демонстрируют, как и Spermophilus parryi и Tamius sibiricus, достаточно распространенную картину геногеографической изменчивости популяций шельфовых островов, когда в последних представлены и наиболее широко распространены те же аллели полиморфных локусов, что и в близлежащих материковых. Несмотря на тысячи поколений раздельного существования явных показателей дифференциации между островными филетическими линиями не выявлено, хотя по отдельным локусам отмечается мозаичный характер распределения аллельных вариантов.
В свою очередь, ареалы M. sikotanensis и M. reх разбиты на изолированные участки, приуроченные к небольшим островам – Кунашир, Шикотан, Ришири, Дайкоку – либо к ограниченным участкам территории больших островов – Сахалин и Хоккайдо. Эти популяции более подвержены действию генетико-автоматических процессов, поэтому накопили значимые генетические различия. Наблюдаемый уровень различий отражает существование раздельных филетических линий, т. е. быструю эволюцию малых популяций.
Предлагается гипотеза последовательных волн миграций, по которой M. sikotanensis и M. rex являются потомками более ранней волны заселения островов. Существуют общие онтогенетические и морфологические характеристики зубов, по которым M. sikotanensis и M. rex выделяются среди остальных Myodes. К таким признакам относятся сроки формирования корневой системы зубов – родовая характеристика, подразделяющая Myodes и Eothenomys Miller, 1896. Как показано Е. Кавамурой (Kawamura, 1988), эволюция полевок Японского архипелага в период средний – поздний плейстоцен шла по пути утраты корнезубости. По данной характеристике эндемичные формы занимают промежуточное положение между современными Myodes и Eothonomys. У них корневая система зубов формируется на поздних стадиях онтогенеза. Возможно, что на примере M.sicotanensis и M.rex мы наблюдаем промежуточную стадию подобного эволюционного процесса у первой волны мигрантов Myodes, тогда как у M. rutilus и
M. rufocanus, заселивших острова в более позднее время, данные эволюционные изменения не обнаруживаются.
Далее в главе проводится сопоставление данных по внутривидовой аллозимной дифференциации М. rutilus, М. rufocanus, M. oeconomus. Минимальный уровень различий характеризует популяции одного и того же острова полевки-экономки. Дифференциация популяций лесных полевок северныхи южных частей о-ва Сахалин несколько выше, а для красной полевки, судя по несмещенным генетическим дистанциям, статистически достоверно выше. У всех видов дифференциация между островными изолятами слегка превышает различия популяций одного острова. Эти превышения достоверны для полевки-экономки, где рассматривались выборки животных из трех бухт о-ва Симушир. Превышения не достоверны для фоновых видов лесных полевок, где рассматриваются популяции о-ва Сахалин. Дифференциация островных изолятов красной и красно-серой полевок несколько выше, чем у полевки-экономки. Эти различия для красной полевки статистически достоверны. Более значимые генетические дистанции характеризуют подвидовой уровень, представленный, с одной стороны, островными выборками, а с другой, пространственно удаленными – материковыми. То есть, как и у большинства материковых видов, в том числе и рассматриваемых в разделе 1 Spermophilus, обнаруживается параллелизм в накоплении морфологических и аллозимных различий в процессе дифференциации. Однако картина может меняться, когда в эволюционный процесс вклинивается хромосомный фактор. Именно такая ситуация рассматривается в следующем разделе диссертации.
Раздел 3. Глава 9. АЛЛОЗИМНАЯ ДИФФЕРЕНЦИАЦИЯ КАК ПОКАЗАТЕЛЬ РОЛИ ХРОМОСОМНОГО ФАКТОРА В ЭВОЛЮЦИОННОЙ ИСТОРИИ ТАКСОНА (НА ПРИМЕРЕ ДАЛЬНЕВОСТОЧНЫХ СЕРЫХ ПОЛЕВОК РОДА MICROTUS Schrank, 1798
Градуалистический тип видообразования предполагает постепенный процесс накопления морфологических, кариологических, молекулярно-генетических поведенческих и других различий, в том числе и по признакам, обеспечивающим репродуктивную изоляцию. На генетическом уровне хорошей иллюстрацией такого процесса являются результаты аллозимных исследований, показывающие увеличение уровня генетической дифференциации от внутривидового к межвидовому уровню. Именно такой тип видообразования широко представлен в рассматриваемых выше Spermophilus и Myodes. В середине прошлого века было предложено понятие внезапного (стасипатрического) или хромосомного видообразования (Воронцов, 1960; White, 1968). Стремительность начальных этапов хромосомного видообразования не предполагает заметных различий других генетических, в том числе аллозимных, характеристик. Накопление значимых аллозимных различий при хромосомном видообразовании может возникать лишь в случае длительного самостоятельного существования разошедшихся форм. Иными словами, соотношение кариологической и аллозимной дифференциации может служить характеристикой эволюционного пути группы. И действительно, среди различающихся хромосомными характеристиками видов-двойников известны пары с минимальными величинами молекулярно-генетических (аллозимных) различий (Фрисман, Павленко, 1987), что соответствует представлению об их недавнем (в эволюционном смысле) возникновении. Однако ситуация неоднозначна, т. к. известны кариотипически различающиеся виды-двойники, обладающие при этом достаточно высоким уровнем молекулярно-генетических различий, что не отрицает их возникновения за счет преобразований кариотипа, но отодвигает в отдаленное прошлое факт его осуществления. Зачастую кариотипические различия обнаруживаются между аллопатрическими таксонами. Если они сопровождаются даже небольшим уровнем различий по другим признакам, то невозможно определить первичность влияния хромосомного либо географического факторов. Так, кариотипические отличия Spermophilus odessanus от S. suslicus, а S. pallidicauda от S. brevicauda могут быть как причиной, так и следствием процесса видообразования, как и обнаруженные нами и описанные в разделе 1 их аллозимные различия.
Тем не менее хромосомное видообразование (по меньшей мере, его начальные этапы) предполагает несостоятельность или недостаточность морфологических характеристик для видовой диагностики и высокую продуктивность методов сравнительной кариологии. Именно такая ситуация наблюдается среди видов серых полевок подрода Microtus. Начиная со второй половины прошлого столетия, на основании кариологических данных в пределах подрода был описан ряд новых видов крайне слабо различающихся морфологически. В составе надвида
M. arvalis Pallas, 1778 обнаружены хромосомно дифференцированные виды-двойники (Мейер и др., 1969) M. arvalis sensu stricto (2n=46) и симпатричный ему на значительной части ареала
M. rossiaemeridionalis Ognev, 1924 (=M. epiroticus Ongrias, 1966; = M. subarvalis Mejer, Orlov, Skholl, 1972 (2n=54)). С территории Муйской котловины в Северном Забайкалье описана муйская полевка (2n=38; NFa=46-50) - M. mujanensis Orlov et Kowalska, 1975 (Орлов, Ковальская, 1978). Из окрестностей озера Эворон в Хабаровском крае описана эворонская полевка (2n=38-40; NFa=51-54) - M. evoronensis Kowalska et Sokolov, 1980 (Ковальская, Соколов, 1980). Лишь опираясь на кариологические данные, удалось поставить точку в дискуссии о видовой обособленности дальневосточной (2n=52; NFa=62) M. fortis Buchner, 1889 и сахалинской (2n=50; NFa=60) M. sachalinensis Vasin, 1955 полевок (Мейер, Волобуев, 1974) и о видовой самостоятельности полевки Максимовича (2n=36-44; NFa=50-60) – M. maximowiсzii Schrenk, 1893. (Ковальская, 1974; Ковальская, 1977; Ковальская и др.,1980; Мейер и др., 1996).
В данной главе на основе сопоставления внутри - и межвидовой аллозимной дифференциации оценивается роль хромосомного фактора в эволюционной истории кариотипически дифференцированных дальневосточных серых полевок.
M. maximowiczii. Полевка Максимовича обитает в припойменных лугах лесной зоны от Забайкалья до Приамурья и северных районов Хабаровского края. Заселяет районы северной Монголии и Северо-Восточного Китая. Данная территория представляет собой область сложной геоморфологической структуры, где ареал вида представляет собой кружевную структуру, включающую серии изолятов и полуизолятов. На российской части ареала выделяются три подвида. В Забайкалье описан M. m. ungurensis Kastschenko, 1913, на территории Амурская обл." href="/text/category/amurskaya_obl_/" rel="bookmark">Амурской обл. и Хабаровского края - M. m. maximowiсzii. M. m. gromovi Vorontsov, Boeskorov, Ljapunova et Revin, 1988 в настоящее время известен выборками из двух географических точек.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
