



 Рисунок 2 – Филогенетическое дерево, построенное на основании фрагмента гена нуклеопротеина (572-776 п. н.) изолятов лиссавирусов, выделенных нами на ЮЗС (выделены жирным шрифтом) со штаммами, представленными в базе данных GenBank
Рисунок 2 – Филогенетическое дерево, построенное на основании фрагмента гена нуклеопротеина (572-776 п. н.) изолятов лиссавирусов, выделенных нами на ЮЗС (выделены жирным шрифтом) со штаммами, представленными в базе данных GenBank
GenBank, показал, что на юге Западной Сибири циркулируют близкородственные генетические варианты вируса бешенства, относящиеся к генотипу 1.
Итогом работы по изучению вируса бешенства явилось подтверждение главной роли диких плотоядных (53,0%) в резервации и распространении рабической инфекции в природных очагах на юге Западной Сибири. Кроме диких плотоядных установлено наличие вируса бешенства у барсуков, косуль, зайцев, сурков, бобров и мышевидных грызунов. Установлено, что периодичность эпизоотий бешенства на юге Западной Сибири в течение 1997–2008 годов составила 3–5 лет. Была установлена зависимость эпизоотологической роли основных резервуаров вируса бешенства от направленности и интенсивности антирабических мероприятий. Увеличение в 3 раза объемов иммунизации домашних животных обеспечило снижение на 46,0% случаев бешенства у этой категории животных. Ежегодная пероральная иммунизация диких лисиц в природных эпизоотических очагах эффективно снижала проявление эпизоотии бешенства на 36,0%. Выявлены фрагменты гена нуклеопротеина лиссавирусов генотипа 1 у насекомоядных летучих мышей, обитающих на территории юга Западной Сибири. Показано близкое генетическое родство лиссавирусов, циркулирующих на территории юга Западной Сибири в популяциях животных разных видов, которые формируют две основные филогенетические группы в пределах генотипа 1.
2.2.2.2. Вирус Западного Нила
Эколого-эпизоотологические характеристики
В настоящей главе диссертационной работы экосистемы юга Западной Сибири были рассмотрены с точки зрения существования в них условий, благоприятных для поддержания циркуляции ВЗН, не связанной с сезонным заносом вируса. При рассмотрении биотических сообществ основное внимание уделялось животным, способным вовлекаться в трансмиссивный цикл ВЗН (птицы, мелкие млекопитающие, кровососущие комары и клещи). Из абиотической компоненты рассматривались физические факторы, влияющие на видовой состав, численность и активность членистоногих переносчиков – температура, влажность и скорость ветра, а также температурный диапазон, определяющий эффективность репликации вируса в их организме. Наличие вирусных маркеров у позвоночных животных и кровососущих членистоногих, участвующих в трансмиссивном цикле ВЗН, а также существование благоприятных условий для репликации вируса свидетельствуют о циркуляции ВЗН в цепочке птица↔комар (клещ)→другое позвоночное животное в пределах определенных биотопов.
Вовлечение в трансмиссивный цикл ВЗН случайных хозяев вируса, не являющихся компонентами природных экосистем – сельскохозяйственных животных, людей – приводящее к развитию заболевания и (или) формированию иммунитета, является косвенным доказательством циркуляции вируса в местных популяциях кровососущих членистоногих и формирования сезонных очагов циркуляции. При составлении характеристики биотопов, выбранных для сбора полевого материала, учитывалось наличие перелетных птиц, мелких млекопитающих и членистоногих кровососущих, способных вовлекаться в трансмиссивный цикл ВЗН, а также антропогенная нагрузка, влияющая на численность и видовой состав животных, участвующих в циркуляции ВЗН. Для большинства выбранных биотопов было характерно присутствие диких перелетных птиц и кровососущих членистоногих, способных вовлекаться в циркуляцию ВЗН. В то же время, биотопы различались по экологическим условиям, влияющим на обитание и активность участников трансмиссивного цикла, а это, в свою очередь, может влиять на циркуляцию вируса в конкретном биотопе.
Подавляющее большинство добытых во всех ландшафтных зонах птиц относились к воробьеобразным, т. е. к ВЗН-восприимчивым. Сравнение распределения вирусных маркеров по сезонам и местам сбора материала выявило определенные различия. При сопоставлении сезонов гг. в Северной Кулунде было отмечено увеличение степени инфицирования птиц в сезоне 2004 г. Степень инфицирования птиц, добытых в южной лесостепи в разные периоды сезона 2004 г. оказалась различной как при сравнении между периодами сборов, так и при сравнении с тем же сезоном в Северной Кулунде. Среди птиц, добытых летом, доля особей, позитивных по наличию одного из главных маркеров ВЗН белка Е, была выше, чем у птиц, добытых весной. В целом, по выявлению гликопротеина и вирусной РНК процент инфицированных птиц в 2004 г. в южной лесостепи был меньше, чем у птиц, добытых в Северной Кулунде. РНК ВЗН не была выявлена ни в одном образце внутренних органов, вирусный белок Е не был обнаружен ни в одном образце головного мозга птиц, добытых в сезоны 2006 и 2007 гг. С большой частотой гликопротеин Е обнаруживался в печени птиц, а в сезоне 2006 г. – в селезенке у одной птицы. Суммарная частота выявления этого маркера составила в сезоне 2006 г. 43,2%, в сезоне 2007 г. – 42,4%.
Отдельно можно отметить особенности распределения гликопротеина Е и РНК ВЗН в разных группах птиц. В группе грачей, добытых в начале мая на юге Барабинской лесостепи, выявление белка Е совпадало с выявлением вирусной РНК, в то время как в головном мозге большей части птиц, добытых там же в летние месяцы, выявлялся гликопротеин Е, а процент птиц с двумя маркерами был в несколько раз меньше. Высокий процент совпадения по обоим маркерам отмечался и у птиц 2004 г. в Северной Кулунде. Большая частота выявления гликопротеина и вирусной РНК у этих птиц свидетельствует о том, что в организме птиц имела место продуктивная ВЗН-инфекция с попаданием вируса в ЦНС и последующей репликацией в головном мозге. В пользу этого говорит тот факт, что часть грачей с положительными находками гликопротеина и РНК ВЗН были найдены мертвыми, хотя нами за время сборов не было отмечено больших эпизоотий среди этих птиц.
Таким образом, по результатам анализа маркеров ВЗН, большинство обследованных птиц как мигрирующих, так и оседлых видов ранее имели контакт с вирусом Западного Нила. Результатом этого у части птиц стало развитие продуктивной ВЗН-инфекции с попаданием вируса в ЦНС, которая могла стать причиной их гибели; у другой части птиц имела место абортивная инфекция, ограниченная внутренними органами. В таком случае, при различном исходе инфекции у птиц, эффективность передачи вируса от них другим участникам трансмиссивного цикла (как позвоночным, так и членистоногим) также должна различаться и коррелировать с распределением в организме этих животных вирусных маркеров. Для проверки этого предположения был проведен анализ внутренних органов млекопитающих на наличие и особенности распределения маркеров ВЗН.
Млекопитающие
Из отловленных мелких млекопитающих в степной зоне большую часть составили мышевидные грызуны. Распределение маркеров ВЗН у зверьков степных и лесостепных биотопов отличалось от такового для подзоны осиново-березовых лесов. Наиболее часто гликопротеин Е и вирусная РНК встречались в головном мозге животных лесостепных биотопов, причем вирусный антиген был обнаружен во всех образцах головного мозга. Почти полное совпадение в выявлении гликопротеина Е и РНК наблюдалось у грызунов, отловленных во влажном колке с действующей колонией грачей, далее по частоте встречаемости маркеров следовали животные, отловленные на остепненном лугу и прибрежной полосе р. Чулым. Этот факт свидетельствует о протекании в организме всех обследованных животных продуктивной ВЗН-инфекции с попаданием вируса в ЦНС.
В Северной Кулунде чаще всего вирусные маркеры встречались у зверьков, отловленных на заросшем поле у оз. Кротово. На втором месте по частоте встречаемости маркеров следовали животные, отловленные на участке полынно-злаковой степи, несмотря на отсутствие в этих биотопах благоприятных условий для активности комаров. Этот факт можно объяснить как перемещением животных в биотопы, где активны комары, так и возможным участием в передаче вируса преимагинальных стадий клещей рода Dermacentor. В целом, частота встречаемости обоих вирусных маркеров (белок Е/РНК) у мелких млекопитающих Северной Кулунды составила 40,0%/12,7%. Таким образом, среди мелких млекопитающих Северной Кулунды доля животных с протеканием продуктивной ВЗН-инфекции заметно меньше по сравнению с животными южной лесостепи.
Кровососущие членистоногие
Комары
Хотя маркеры ВЗН у комаров нами выявлены не были, в Северной Кулунде и на юге Барабинской лесостепи имеются виды комаров, способные участвовать в трансмиссивном цикле ВЗН и для которых характерна высокая численность в некоторых биотопах. Это создает предпосылки для поддержания циркуляции вируса в период их активности.
Клещи
Результат анализа на распределение маркеров ВЗН у иксодовых клещей из разных биотопов в сезоне 2006 г. показали, что наиболее часто белок Е и РНК ВЗН встречались у имаго Ix. рersulcatus и Ix. pavlovskyi с участием всех возрастных стадий.
Молекулярно-биологические характеристики вариантов ВЗН
Из части образцов, в которых были выявлены вирусные маркеры, с помощью ОТ-ПЦР были получены фрагменты кДНК, соответствующие участку гена белка Е ВЗН, которые были секвенированы. Секвенированные образцы имели следующее происхождение: два образца от серых ворон из Северной Кулунды 2003 г. (ChCorvus10-03, ChCorvus31-03), три образца от грачей из южной лесостепи 2004 г. (ChRook7-04, ChRook14-04, ChRook27-04), образец от красной полёвки из южной лесостепи 2004 г. (ChRodent-04), два образца от заболевших людей (Пациенты №1 и №3, ChiWS-04 и HumWS-04 соответственно) и два пула имаго клещей Ix. persulcatus и Ix. pavlovskiy из подзоны осиново-березовых лесов 2006 г. (Tomsk-1, Tomsk-2).
Генотипирование исследуемых нуклеотидных последовательностей позволило идентифицировать эти последовательности как относящиеся к генотипу Ia вируса Западного Нила. Западносибирские варианты ВЗН гг. и 2006 г. оказались высоко гомологичными (не более 1-3% расхождения) с волгоградским штаммом VLG-4 ВЗН. При анализе филогенетического дерева (рис.3) можно видеть, что исследованные варианты характеризуются невысоким уровнем генетических отличий. Можно также видеть, что исследованные западносибирские варианты ВЗН не имеют гостальной специфичности. Филогенетический анализ показал, что выявленные в гг. и 2006 г. последовательности ВЗН кластеризуются вместе с другими современными российскими изолятами ВЗН. Это говорит о том, что эти варианты вируса генетически связаны со штаммами ВЗН, изолированными на юге Европейской части России. В отношении вариантов, выделенных от птиц, полученные результаты согласуются с орнитологическими данными, и это может служить доказательством заноса ВЗН перелётными птицами на юг Западной Сибири с эндемичных территорий на юге РФ. Поскольку для клещей Ix. persulcatus и особенно Ix. pavlovskiy, птицы являются прокормителями, выявление вариантов ВЗН в клещах свидетельствует об участии птиц в заносе вируса и передаче членистоногим переносчикам. Генетическая близость исследуемых вариантов с патогенными для людей штаммами ВЗН юга России позволяет предполагать, что западносибирские варианты ВЗН обладают эпидемическим потенциалом и могут быть причиной заболевания ВЗН людей на юге Западной Сибири.
Из филогенетического дерева можно видеть, что варианты ВЗН от серых ворон и грача (ChRook27-04) образуют одну группу. По сравнению с прототипным штаммом VLG-4, они характеризуются общей консервативной заменой Arg94®Lys, которая также свойственна вариантам ВЗН, выделенным в различных регионах мира и относящимся к генотипам Ib, II, III и IV, но не выявлена у чирков и грачей юга Западной Сибири в 2002 году (генотип Ia). Каждый вариант от серых ворон имеет индивидуальные консервативные уникальные замены – у одного обнаружена Ala73®Thr, у другого – Pro89®Ala, также не встречавшиеся ранее у других вариантов вируса. В целом эта группа характеризуется консервативной уникальной заменой Pro76®Ala и значимой заменой Leu69®Hys, выявленной у грача и вороны, также являющейся уникальной. Эти замены выявлены в вариантах ВЗН от птиц, принадлежащих к видам с разным миграционным статусом, добытых в разное время и в разных ландшафтных зонах, что может указывать на общий источник инфицирования птиц ВЗН, а также на возможное участие организма хозяина в воздействии на изменчивость вируса.
Таким образом, генотипирование позволило характеризовать западносибирские варианты ВЗН как генетически близкие между собой и патогенным для людей штаммам ВЗН, изолированным на юге России. Сравнительный аминокислотный анализ выявил замены только в вариантах от восприимчивых к ВЗН врановых птиц, несмотря на значительную генетическую близость этих вариантов ВЗН с вариантами от других хозяев. Полученные результаты свидетельствуют о способности вируса адаптироваться к разным хозяевам в различных ландшафтно-экологических условиях, а также об эпидемическом потенциале ВЗН, циркулирующего на юге Западной Сибири.
Итогом по изучению циркуляции вируса западного Нила на территории юга Западной Сибири явилось выявление циркуляции ВЗН среди воробьеобразных птиц (Passeriformes), мигрирующих и оседлых видов, в степной, лесостепной зонах и подзоне осиново-березовых лесов юга Западной Сибири. Также было установлено, что в циркуляцию ВЗН в степной, лесостепной зонах и подзоне осиново-берёзовых лесов юга Западной Сибири вовлекаются мелкие млекопитающие – грызуны, насекомоядные и мелкие хищники. Среди комаров-эудоминантов выявлены эпидемиологически значимые виды, способные принимать участие в трансмиссивном цикле ВЗН. Показано вовлечение в циркуляцию ВЗН имагинальных и преимагинальных стадий иксодовых клещей Ix. persulcatus и Ix. pavlovskiy в подзоне осиново-берёзовых лесов юга Западной Сибири. В степной, лесостепной зонах и подзоне осиново-берёзовых лесов юга Западной Сибири показано формирование природных очагов ВЗН, не связанных с сезонным заносом. Было установлено вовлечение в трансмиссивный цикл ВЗН на юге Западной Сибири случайных участников – сельскохозяйственных животных и людей.
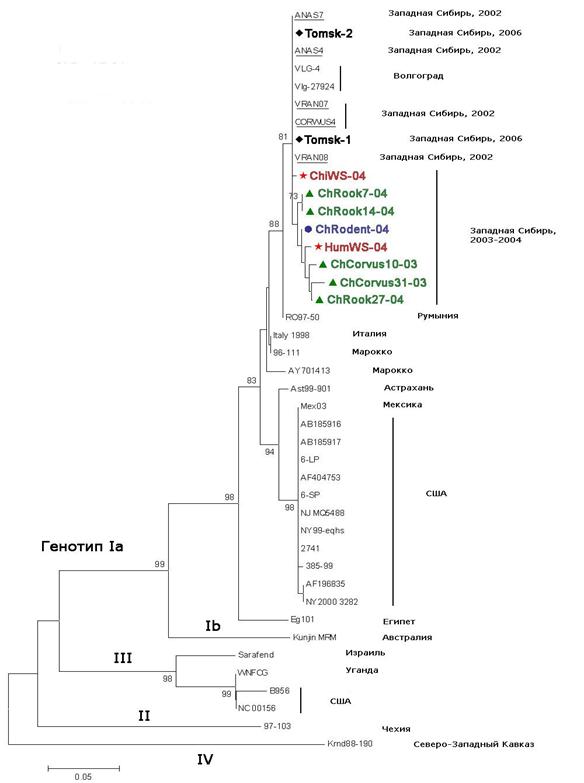
Рисунок 3 – Филогенетическое дерево полевых изолятов и штаммов ВЗН, построенное на основании сравнения нуклеотидных последовательностей фрагмента гена белка Е (322 нуклеотида) со штаммами, представленными в базе данных GenBank.
Примечания: – птицы (серые вороны и грачи); – красная полёвка; - - люди; – клещи Ix. persulcatus и Ix. Pavlovskiy (фрагменты РНК выявленные нами у различных хозяев на ЮЗС).
Секвенирование и последующий анализ фрагментов гена белка Е ВЗН, выделенного от разных хозяев, показал, что западносибирские варианты ВЗН принадлежат генотипу Iа ВЗН, являются генетически близкими между собой и близки с патогенными для людей штаммами ВЗН, изолированными на юге России.
2.2.2.3. Вирус гриппа
Эколого-эпизоотологические характеристики
Анализ орнитологических данных показал, что юг Западной Сибири является уникальным местом, где пересекаются три из шести миграционных путей, проходящих через территорию Евразии. Через лесостепную и степную зону юга Западной Сибири мигрирует более 250-ти видов птиц, причем на отдельных ее участках весной может обитать до 220 видов птиц одновременно. Массовые миграции птиц протекают с конца марта до первой половины июня и со второй половины июля почти до середины октября. В послегнездовое время и при пролете образуются локальные скопления с включением до 50 видов птиц с общей численностью до 20 и более тысяч особей единовременно. При массовых скоплениях возникают контакты птиц, посещающих в течение своего жизненного цикла различные ландшафтные зоны ряда регионов и континентов (от Западной Сибири до субтропиков и тропиков Европы, Азии и Африки). Все это создает благоприятные условия для распространения возбудителей различных болезней, в том числе вируса гриппа птиц. Основными объектами при сборе проб для лабораторного анализа являлись водоплавающие и околоводные птицы: гуси, утки, кулики, чайки и крачки и др. Кроме этого, собирали материал от птиц синантропных видов: голубей, сорок, ворон и воробьев. За период с 2002 по сентябрь 2009 года в обследуемом районе было проведено 109 экспедиций по сбору проб от диких и домашних птиц. Было собрано и обследовано на наличие вируса гриппа 19856 проб. За время наблюдения (исключая период эпизоотии высокопатогенного вируса гриппа H5N1 годов) нами было выделено 219 изолятов вируса гриппа от 34 различных видов диких птиц.
По результатам анализа, вирусы гриппа А, циркулировавшие в предэпизотический период ( гг). среди диких птиц на юге Западной Сибири, оказались филогенетически близкими вирусам гриппа, изолированным от диких птиц в странах Европы и южной Азии. Это указывает на распространение перелетными птицами различных вариантов вирусов гриппа А между различными регионами Евразии.
Эпизоотия высокопатогенного вируса гриппа Н5 на юге Западной Сибири
и ее последствия
Первая в России эпизоотия высокопатогенного вируса гриппа H5N1 субтипа началась на юге Западной Сибири в Новосибирской области в июле 2005 года. Первичные очаги гриппа птиц представляли собой водоемы, находящиеся на территории населенных пунктов, либо в непосредственной близости от них. Пик активности возбудителя гриппа птиц совпал с перемещением выводков диких птиц с мест гнездования на кормовые водоемы с наименьшим фактором беспокойства, которыми, как правило, и являются озера близ населенных пунктов. Развитие эпизоотии в августе, сентябре можно связать с тем, что инфицированные домашние утки и гуси занесли вирус во дворы, где происходила его передача другим видам домашней птицы и, в первую очередь, наиболее чувствительным: курам и индюкам.
Первый материал от погибшей птицы поступил на исследование 20 июля 2005 года из села Суздалка (Новосибирская область), где была зарегистрирована гибель домашних птиц. Вирусы гриппа Н5N1 были изолированы из органов трех погибших птиц: одной индюшки и двух кур. Гены, кодирующие гемагглютинин у выделенных изолятов, были секвенированы. Их филогенетический анализ показал, что выделенные штаммы образуют кластер (индекс поддержки = 99) с H5N1 штаммами, изолированными от птиц во время вспышки птичьего гриппа на озере Цинхай (Qinghai) в Китае в апреле 2005 года. У всех выделенных нами изолятов сайт протеолитического расщепления был – ERRRKKR/GL, что соответствовало сайту расщепления высокопатогенных штаммов, циркулирующих в странах Юго-Восточной Азии. Тест на патогенность с использованием 6-ти недельных цыплят также подтвердил, что все выделенные штаммы являются высокопатогенными для птиц (HPAI).
Таким образом, нами впервые было показано, что массовая гибель домашней птицы на юге Западной Сибири летом 2005 года была вызвана высокопатогенным вирусом гриппа Н5N1 субтипа, генетически родственным вирусам, выделенным во время майской вспышки (2005г.) на озере Цинхай (Северный Китай).
Всего за время эпизоотии 2005 года нами было собрано или получено 4993 образца от диких и домашних птиц и выделено 55 изолятов высокопатогенного вируса гриппа H5N1 субтипа.
Исследование роли синантропных птиц в эпизоотии высокопатогенного гриппа
у диких и домашних птиц на юге Западной Сибири
При изучении эпизоотии и особенно ситуации в постэпизоотический период важным оставался вопрос о роли синантропных птиц в распространении высокопатогенного вируса гриппа птиц и возможности формирования локальных очагов данного патогена у птиц, не являющихся основным резервуаром. Для этого был проведен мониторинг вируса гриппа А среди синантропных видов птиц в эпизоотический и постэпизоотический периоды (с сентября 2005 по март 2006) в местах первичного распространения гриппозной инфекции на юге Западной Сибири. Было собрано 458 образцов от 10 видов птиц, в т. ч. 70 образцов в эпизоотический и 388 – в постэпизоотический периоды. В результате проведенных исследований ни одного высокопатогенного вируса гриппа H5N1-субтипа выделено не было. Был выявлено один изолят непатогенного вируса гриппа субтипа H4N6 у обычной сороки. Полученные данные указывают на то, что синантропные птицы играли незначительную роль в поддержании циркуляции высокопатогенного вируса гриппа H5N1 во время эпизоотии гг. на юге Западной Сибири.
Особенности циркуляции вируса гриппа у диких и домашних птиц
в постэпизоотический период
Несмотря на то, что осенью 2005 и весной 2006 годов на юге Западной Сибири было зафиксировано несколько случаев локальной гибели домашней птицы, мы считаем, что эпизоотия высокопатогенного гриппа Н5N1 на этой территории закончилась осенью 2005 года с окончанием осенней миграции птиц и наступлением холодов. Анализируя циркуляцию вирусов гриппа у диких и домашних птиц в летне-осенний период 2005 года (во время эпизоотии), хотелось бы отметить, что мы не выявили циркуляции ни одного иного серотипа вируса гриппа, кроме высокопатогенного Н5N1. Весна 2006 года характеризовалась также практически полным отсутствием любых субтипов вируса гриппа у диких птиц обследованных на территории юга Западной Сибири. Был выделен только один изолят вируса гриппа Н5N1 из 237 собранных проб. Характерно, что утка была отстрелена во время весенней охоты и была клинически здоровой. Этот факт указывал на то, что во время весенней миграции 2006 был возможен повторный занос вирусов Н5N1 субтипа на юге Западной Сибири, что в дальнейшем и случилось. Так, в мае 2006 года одновременно в двух селах Омской и Новосибирской областей была зафиксирована гибель домашней птицы на частных подворьях. Из органов погибших птиц был выделен и охарактеризован высокопатогенный вирус гриппа Н5N1 субтипа, генетически близкий вирусам, циркулировавшим на территории Западной Сибири в 2005 году. Все вспышки, описанные на этот период, были вызваны вирусом гриппа H5N1-субтипа, принадлежащего к генетической линии Цинхай-подобных вирусов – клада 2.2 (clade 2.2) (рис. 4, 5) .
При проведении мониторинга вируса гриппа на юге Западной Сибири в последующие годы () нами было собранно и обследовано 14307 проб от диких и домашних птиц. За этот период было выделено 133 изолята вируса гриппа различного субтипа, не считая высокопатогенных H5N1. Анализ полученных данных показал, что начиная со второй половины 2006 года, появляется разнообразие выделяемых субтипов вируса гриппа, которое полностью отсутствовало во время эпизоотии 2005 года и в постэпизоотический период 2006 года (январь – август). Во второй половине 2006 года и в 2007 году были выделены и субтипы H3N8, H5N2, H4N6 и др. Аналогичная картина наблюдалась и в 2008 и 2009 годах.
При обследовании колонии сизых чаек (Larus canus) в июле 2006 г. на оз. Чаны в Новосибирской области из клоакального смыва, взятого прижизненно, был выделен изолят H5N1. Выделение высокопатогенного вируса гриппа H5N1-субтипа от клинически здоровой особи является чрезвычайно интересным и важным в эпизоотологии данного заболевания. Данный факт указывает на то, что в популяциях некоторых видов птиц наступило равновесие между паразитом и хозяином, когда паразиту (в данном случае вирусу гриппа) не целесообразно уничтожать своего хозяина, а предпочтительно сосуществовать вместе.
В мае–июне 2009 г. новая вспышка высокопатогенного гриппа птиц была отмечена на берегу оз. Увс-Нуур. Филогенетический анализ нуклеотидных последовательностей гена гемагглютинина выделенных изолятов показал, что все они принадлежат к кладе 2.3.2 (рис. 4, 5) и наиболее близки вирусам, выделенным ранее в Юго-Восточной Азии и Китае. Судя по всему, на территорию России вирусы с новым генотипом были занесены перелетными птицами таким же образом, как и в гг.
Молекулярно-биологические характеристики вариантов вируса гриппа,
выделенных на юге Западной Сибири
Филогенетический анализ позволяет определить и показать степень родства вирусов или отдельных генов вирусов, циркулирующих на разных территориях. В связи с этим для нас представлял большой интерес анализ не только орнитологических данных о разлете птиц с территории юга Западной Сибири, но и родство вирусов гриппа и/или генов вирусов, выделенных в различных регионах. Учитывая, что при мониторинге вируса гриппа у диких птиц в основном выделялись и определялись вирусы Н2, Н3 и Н4 субтипов гемагглютинина, нами был проведен филогенетический анализ генов, кодирующих гемагглютинин вируса гриппа именно этих субтипов.
Результаты филогенетического анализа показали, что вирусы гриппа подтипа Н4, изолированные нами в период гг. от диких уток, отстреленных на юге Западной Сибири, эволюционно близки вирусам, изолированным от диких уток годом ранее в Европе; изоляты вируса гриппа подтипа Н2, выделенные нами в 2003 году от диких уток на юге Западной Сибири, эволюционно близки штаммам, изолированным от диких уток двумя годами ранее в южной Азии и штамму, изолированному от дикой утки 4-мя годами ранее в Европе; изоляты гриппа А подтипа Н3, циркулировавшие среди диких птиц на юге Западной Сибири, эволюционно близки вирусам, циркулировавшим практически в то же время среди диких птиц в Европе и штамму, изолированному ранее в Китае.
Таким образом, вирусы гриппа А различных субтипов, циркулировавшие среди диких птиц на юге Западной Сибири, филогенетически близки вирусам гриппа, изолированным от диких птиц в эти же годы в странах Европы и южной Азии. Это указывает на широкие связи юга Западной Сибири с различными регионами Евразии и подтверждает орнитологические данные о важной роли юга Западной Сибири в быстром распространении различных вариантов вирусов гриппа А во время миграций птиц на этом континенте. Быстрое распространение родственных вариантов вирусов гриппа птиц между районом проведения мониторинга и значительно от него удаленными регионами Евразии, установленное нами при орнитологическом и филогенетическом анализе изолятов вируса гриппа, объясняется уникальным пересечением миграционных путей диких птиц на юге Западной Сибири. Во время весенних миграций в данный регион могут быть занесены вирусы гриппа, циркулирующие как среди птиц восточно-азиатского миграционного русла, пролегающего через Дальний Восток и Юго-Восточную Азию, так и среди птиц восточно-европейского миграционного потока.
Изучение высокопатогенных вирусов гриппа, выделенных на территории
юга Западной Сибири
Биологические свойства штаммов вируса гриппа H5N1-субтипа,
выделенных от диких и домашних птиц на юге Западной Сибири ( гг.)
Учитывая большую экономическую и эпизоотическую значимость высокопатогенных вирусов гриппа H5N1, изучению биологических свойств данной группы вирусов было уделено особое внимание. Культивирование в системе развивающихся куриных эмбрионов показало, что все изученные нами штаммы вируса гриппа H5N1 вызывали гибель развивающихся куриных эмбрионов в течении 2 суток культивирования. В процессе визуального анализа инфицированных эмбрионов регистрировались поверхностные геморрагии и образование желеобразной аллантоисной жидкости. Наблюдалось глубокое поражение зародышевых оболочек эмбриона. Для определения оптимальной дозы заражения, являющейся интегральной характеристикой, описывающей инфекционные свойства различных штаммов вируса, был проведен подбор условий культивирования и оптимизирована доза заражения. Все штаммы вызывали высокую продукцию вируса в РКЭ.
Логическим продолжением работ на эмбрионах явилось изучение экспериментальной инфекции гриппа, вызываемого штаммами H5N1, у кур как наиболее релевантной модели заболевания у птиц. Тест, который был использован нами для количественной оценки патогенности конкретного штамма, заключался в определении величины индекса IVPI (intravenous pathogenicity index) при внутривенном заражении кур. Индекс IVPI может принимать значение от 0.00 до 3.00. Принято, что при значении IVPI=1.2 и выше штамм считается высокопатогенным, в остальных случаях – низкопатогенным. Первые клинические проявления инфекции и начало гибели птиц были отмечены на 1-2-е сутки после инфицирования. К третьим суткам для всех изученных штаммов, кроме A/common gull/Chany/P/2006, падеж составил 100%. Типичная картина заболевания выглядела следующим образом: снижалась двигательная активность; нахохлившись, птицы сбивались в кучки; у отдельных особей регистрировались признаки поражения желудочно-кишечного тракта. Далее отмечались поражения нервной системы: изменение походки, больная птица при передвижении приседала, прихрамывала, падала, упиралась клювом в пол, вытягивала ноги, крылья, прикрывала глаза, либо падала на спину и не могла перевернуться, беспокойно подергивала конечностями. При внешнем осмотре погибшие птицы лежали на боку с вытянутыми конечностями, либо на спине, чаще голова запрокинута, наблюдались бледность, либо умеренная цианотичность гребня и сережек. При патологоанатомическом исследовании выявлены: бледность, либо умеренная цианотичность мышечной ткани; легкие отечные, наполненные кровью, с геморрагиями и очагами некроза, тонут в воде, на разрезе темно-бордового цвета, с поверхности разреза выступает темно-бордовая, не свернувшаяся кровь. Сосуды брыжейки желудочно-кишечного тракта инъецированы, зоб и мышечный желудок наполнены кормовыми массами, либо вздуты. Печень темно-вишневого или кирпичного цвета. Желчный пузырь наполнен. Почки отечные, наполненные кровью, дряблые, имеющие зернистую консистенцию на разрезе, структура почек не различима. Селезенка темно-вишневого или темно-розового цвета с синюшным оттенком.
В результате проведенного исследования было показано, что все штаммы вируса гриппа птиц H5N1-субтипа, выделенные на юге Западной Сибири, были высокопатогенными для кур.
Поскольку ВОЗ рассматривает вирус H5N1-субтипа как потенциально пандемичный, актуально изучение патогенности различных вариантов вируса гриппа H5N1 для млекопитающих. Общепризнанной моделью изучения гриппозной инфекции является мышиная модель. Для оценки восприимчивости и летальности мышей было проведено интраназальное заражение их рядом штаммов, выделенных от диких птиц в разные годы (табл. 1).
Таблица 1 – Штаммы вируса гриппа, использованные в эксперименте по заражению мышей
Штамм | Lg ГАЕ/мл | Lg EID50/мл | LD50/мл | |
1 | A/duck/Omsk/1822/2006 | 3,41±0,15 | 8,38±0,23 | 6,5±0,18 |
2 | A/Black-headed gull/Tyva/115/09 | 3,71±0,15 | 9,0±0,20 | 6,4±0,25 |
3 | A/duck/Tuva/01/06 | 3,71±0,15 | 9,13±0,34 | 7,0±0,26 |
4 | A/common gull/Chany/P/2006 | 4,61±0,15 | 9,75±0,30 | 7,3±0,22 |
Примечание: В таблице приведены средние значения показателя (M±m), p<0.05; доверительный интервал определения гемагглютинирующей активности при двукратных разведениях аллантоисной жидкости 0.30 lgГАЕ/мл.
Все исследованные штаммы вызывали острую летальную инфекцию у белых мышей при интраназальном заражении, иными словами, оказались высокопатогенными для мышей. Таким образом, все выделенные штаммы потенциально опасны для млекопитающих.
Молекулярно-биологический и филогенетический анализ генов HA и NA штаммов
вируса гриппа H5N1-субтипа, выделенных на юге Западной Сибири
от диких и домашних птиц ( гг.)
Как уже говорилось выше, важной характеристикой вируса является первичная структура его генома. Для вируса гриппа А актуальны исследование и молекулярно-биологический анализ структуры его поверхностных белков – гемагглютинина HA и нейраминидазы NA. Эти белки являются важнейшими антигенами вируса и непосредственно участвуют в инфекционном процессе. Филогенетический анализ показал, что все выделенные на юге Западной Сибири в гг. штаммы относятся к двум генетическим группам, в соответствии с последовательностью гена HA. Это клады 2.2 и 2.3.2. (рис. 4, 5). При этом штаммы, выделенные на юге Западной Сибири во время первых эпизоотий гриппа птиц, образуют отдельный кластер и принадлежат к Цинхай-подобным штаммам (клада 2.2) (рис. 4, 5). Штаммы A/Black-headed gull/Tyva/115/09, A/Great crested grebe/Tyva/120/09, A/Little grebe/Tyva/118/09 и A/Spoonbill/Tyva/117/09 принадлежат к совершенно другой кладе – 2.3.2. (Гуанкси/Фуджан-подобные штаммы) (рис. 4, 5). Генетически они наиболее близки штаммам, выделенным в Японии в гг., и штаммам, вызвавшим вспышку среди диких и домашних птиц в Приморском крае в апреле 2008 г.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
