

ПРАКТИЧЕСКИЕ ЗАНЯТИЯ
Тема 11. РАННЕЕ РАЗВИТИЕ ПЛАЦЕНТАРНЫХ МЛЕКОПИТАЮЩИХ
Цель занятия - ознакомиться с особенностями развития Плацентарных млекопитающих.
Прежде всего, обращает на себя внимание микроскопически малая величина яиц, которые в связи с утратой желтка характеризуются как олиголецитальные или даже алецитальные. Переход к внутриутробному развитию повлек за собой и упрощение в строении яйцевых оболочек: у большинства млекопитающих отсутствуют третичные оболочки и сохраняется лишь тонкая прозрачная оболочка (zona pellucida). В связи с утратой желтка развитие зародышей млекопитающих идет по голобластическому типу. С самого начала дробление характеризуется рядом особенностей: оно происходит медленно (цикл 18-20 часов) и асинхронно. Расположение бластомеров, по крайней мере в период первых митотических циклов, имеет уникальную ориентацию (ротационное); уже со второй половины второго цикла начинается экспрессия некоторых генов зиготы.
После 8-клеточной стадии, во время следующих делений дробления формируется морула. В это время в концептусе совершаются два важных процесса компактизация и поляризация бластомеров. У мышей в конце 8-клеточной стадии бластомеры в результате более тесного прилегания друг к другу изменяют свою форму от сферической до уплощённой, при этом контур концептуса сглаживается. Между уплощающимися клетками, расположенными на поверхности 16-32 клеточной морулы, возникают плотные контакты; и этот слой изолирует внутренние клетки округлой формы, связанные между собой щелевыми контактами.
Первоначально морула не имеет полости, однако в ходе процесса, который называется кавитацией, наружные клетки начинают «накачивать» жидкость внутрь морулы, что вызывает образование полости. При этом центральные клетки оттесняются к одному из полюсов полости в виде компактного агрегата клеток – эмбриобласта или внутренней клеточной массы (ВКМ). Так формируется особая стадия развития млекопитающих, называемая бластоцистой. Концептус на этой стадии представляет собой асимметричный пузырёк, наружная однослойная стенка которого выполнена клетками трофобласта; на одном из полюсов к трофобласту изнутри плотно примыкает ВКМ. В сформированной бластоцисте принято различать муральный трофобласт, образующий стенку полости, и полярный трофобласт, покрывающий эмбриобласт. Трофобласт в последующем даст разнообразные исключительно внезародышевые структуры, активно участвующие в имплантации зародыша в эндометрий матки и составляющие плодную часть плаценты. Эмбриобласт (или зародышевый узелок) состоит из тотипотентных клеток, представляющих не только эмбриональные стволовые линии зародыша, но и дающих материал некоторых внезародышевых структур. Эмбрионально-абэмбриональная ось в бластоцисте перпендикулярна анимально - вегетативной оси.
Происхождение от ранних Рептилий, сохранивших черты организации Амфибий, и вторичная утрата желтка в связи с переходом к внутриутробному развитию и матротрофному питанию зародыша и плода, причудливым образом преломляются в ключевых морфогенетических процессах и, прежде всего, в гаструляции. Вскоре после образования бластоцисты (у мыши через 4,5 суток после оплодотворения) на поверхности ВКМ, обращённой в полость дифференцируется третья линия клеток – гипобласт или первичная эндодерма. Эти клетки покидают зародышевый узелок либо путём ингрессии, либо в результате деляминации и распространяются по внутренней поверхности мурального трофобласта. Эта популяция клеток производит только внезародышевую эндодерму, проксимальный (висцеральный) и дистальный (париетальный) отделы которой формируют впоследствии внутреннюю выстилку желточного мешка. После выделения гипобласта, оставшаяся часть ВКМ называется эпибластом и включает линии клеток зародыша и клеток, дающих внезародышевую мезодерму. С этого момента небольшое щелевидное пространство между эпибластом и гипобластом можно по аналогии с такой же полостью у птиц обозначить как бластоцель, а стадию, на которой происходят описанные события – бластулой млекопитающих.
Помимо присутствия очень сложно организованных провизорных образований зародыш млекопитающих отличается от эмбрионов других позвоночных тем состоянием, в котором он к моменту гаструляции. Это, прежде всего, касается размеров зародышей к началу гаструляции. У млекопитающих гаструляция начинается, когда эпибласт зародыша, например, мыши насчитывает всего 600 клеток. Зародыш шпорцевой лягушки на сравнимой стадии имеет несколько тысяч клеток. По-видимому, именно это обстоятельство является причиной того, что эмбрион млекопитающих начинает расти в очень высоком темпе в предимплантационном периоде.
Гаструляция у всех млекопитающих совершается по такому же сценарию, что и эмбрионов птиц, т. е. через образование и функционирование зародышевой полоски. Образованию зародышевой полоски (ЗП) предшествует либо распластывание клеток эпибласта в виде зародышевого диска (как это происходит у копытных, кроликов, приматов и человека), либо его разрастание в дистальном направлении вдоль длинной оси имплантационной крипты матки с последующим появлением в нём центральной полости и формированием так называемого яйцевого цилиндра (как это имеет место у мышевидных грызунов). Но в обоих случаях клетки эпибласта в одной точке на краю гомогенного эпителия, начинают продуцировать мезодерму и формируют ЗП. Эта точка определяет задний полюс будущего эмбриона. Таким образом, диаметрально противоположная сторона диска или цилиндра теперь с полным основанием может рассматриваться как передний полюс зародыша. Полоска с момента своего появления начинает удлиняться от точки своего зарождения на ⅔ длины area pellucida или до дистальной вершины яйцевого цилиндра. Здесь также, как и у зародыша курицы, образуется специальная структура, называемая «узелком». Узелок – образование, гомологичное «организатору» эмбрионов других позвоночных. Выселяющиеся через узелок клетки дают осевую мезэндодерму, которая содержит в своем составе собственно мезодерму, сосредоточенную по средней линии эмбриона (прехордальную пластинку и хорду) и дефинитивную эндодерму кишечной трубки. Клетки аксиальной мезодермы (хордомезодермы) на этих стадиях отличаются от других клеток существенно меньшими размерами. До момента образования узелка и связанного с этим появления кишечной эндодермы эпибласт на всём своем протяжении подостлан гипобластом (висцеральной эндодермой). Задний конец ЗП продуцирует внезародышевую мезодерму, тогда как материал латеральной пластинки и параксиальной мезодермы выселяется из среднего участка ЗП.
Таким образом, образование зародышевой полоски является действительно критическим событием в детерминации не только переднезадней, но также и дорсовентральной оси тела. А поскольку ориентация лево-правой оси зависит от направления двух предыдущих осей, то паттерн лево-правой организации, в конечном счёте, также связан с формированием зародышевой полоски.
На стадии бластулы или в начале гаструляции концептус попадает из яйцевода в матку и здесь имплантируется, т. е. внедряется целиком (например, у приматов) или только выступающими разрастаниями трофобласта (эктоплацентарный конус у мышевидных грызунов) в слизистую оболочку матки, вступая с ней в тесные обменные взаимоотношения. У некоторых млекопитающих участок трофобласта над зародышевым диском редуцируется, так что наружный слой зародышевого диска на стадии зародышевого щитка, ранее покрытый трофобластом, теперь оказывается снаружи и своими краями смыкается с уцелевшими частями трофобласта в один непрерывный слой. Отпадающий участок трофобласта называется рауберовым слоем. У других форм полярный трофобласт не отпадает, а только разрывается, обеспечивая, тем самым, разворачивание ставшего к этому времени полым зародышевого узелка в зародышевый диск. При этом эпибласт (а позже, эктодерма) зародыша составляет непрерывный слой с трофобластом. Это обстоятельство позволяет рассматривать трофобласт образованием, гомологичным эктодерме серозной оболочки (хориону) птиц. С этого момента трофобласт принято называть трофэктодермой.
Инвазия трофобласта в слизистую оболочку матки вначале происходит с помощью простых выростов, состоящих только из слоя трофэктодермы. Такие выросты называются первичными ворсинками. Позднее, когда в ворсинки проникает мезенхима, образующая в них кровеносные сосуды, они называются вторичными ворсинками. Трофэктодерма, вместе с подстилающей её мезенхимой и сосудами составляет хорион - плодный элемент плаценты, и является эволюционным усложнением серозной оболочки пресмыкающихся и птиц.
Характерно, что, несмотря на отсутствие желтка у плацентарных млекопитающих, тем не менее, имеется желточный мешок. Желточный мешок у них образуется подобно тому, как это происходит у других амниот, и представляет собой ту часть первичной кишки, которая не включается в тело зародыша, когда обособляется при образовании амниотических складок. В его образовании принимает участие гипобласт (внезародышевая эндодерма) и висцеральный листок внезародышевой мезодермы по мере его прорастания в пространство между гипобластом и трофобластом. Здесь клетки мезодермы образуют густую сеть кровеносных сосудов желточного круга кровообращения. Первое время (особенно в развитии копытных млекопитающих) эти сосуды обеспечивают газообмен и питание зародышей, поскольку через выросты трофэктодермы они связаны со слизистой оболочкой матки. Таким образом, хотя желточный мешок у млекопитающих и не выполняет более роли структуры, ассимилирующей питательные вещества желтка, однако, за ним сохраняются важные функции органа первичного кроветворения, через него осуществляется транслокация первичных половых клеток к месту их назначения, кроме того, через сосудистую систему желточного мешка какое-то время совершается газообмен и питание. Правда эта новая функция носит временный характер и позднее переходит к аллантоису и хориону.
Несмотря на более чем надёжную защиту, которую получает зародыш млекопитающих в утробе матери, над ним также развивается дополнительная оболочка – амнион – представляющая часть слоя соматоплевры. У разных видов млекопитающих амнион формируется по-разному. У кролика, копытных и хищных амнион возникает, как у рептилий и птиц, в форме амниотических складок соматоплевры, надвигающихся на тело эмбриона со всех сторон. У рукокрылых (летучие мыши), некоторых насекомоядных и у приматов еще на стадии зародышевого узелка часть его клеток в центре расходятся, образуя небольшую полость (схизоцельный способ). Узелок приобретает вид пузырька, верхняя стенка которого предстает как эктодерма амниона, его полость является амниотической полостью, а утолщенное дно пузырька является эпибластом, в котором вскоре будет формироваться ЗП. Внезародышевая мезодерма, выселяющаяся из каудального отдела ЗП, будет распространяться в разных направлениях, в том числе и по эктодерме амниона, формируя его второй клеточный слой.
Когда внезародышевая мезодерма расщепляется, и ее соматический листок соединяется с трофэктодермой, слой, ставший теперь внезародышевой соматоплеврой млекопитающих, называют хорионом. После образования амниона хорион полностью окружает зародыш со всеми его провизорными образованиями и является поверхностью раздела между тканями зародыша и матки. Если у птиц основная функция хориона – газообмен, то у млекопитающих его назначение разнообразнее; это, в первую очередь, перенос питательных веществ и удаление продуктов обмена, но в то же время и разнообразные синтезы, а также секреция активных регуляторных факторов, например, гормонов.
Аллантоис у млекопитающих образуется в виде дивертикула задней кишки вскоре после её образования (а у мышевидных грызунов ещё раньше, уже на стадии головного отростка). Его стенка образована спланхноплеврой (эндодерма + висцеральный листок мезодермы). По мере роста аллантоиса участок вблизи задней кишки сужается и превращается в цилиндрическую ножку, тогда как его дистальная часть пузыревидно расширяется. В результате периферического роста аллантоис примыкает к хориону и сливается с ним мезодермальными слоями, превращаясь в т. н. хориоаллантоис, обильно снабжённый кровеносными сосудами. Функции аллантоиса у млекопитающих претерпели некоторую коррекцию, причём у разных видов в разной степени. Функция накопления и нейтрализации продуктов азотистого обмена (мочевины и мочевой кислоты), имеющая первостепенное значение в развитии зауропсид, у некоторых млекопитающих (копытные) сохраняется, но на относительно непродолжительном отрезке беременности, у других млекопитающих, с быстрым развитием плаценты, эта функция очень рано становится неактуальной. Например, у приматов и у человека аллантоис, как таковой, имеет зачаточный характер; его эндодермальная часть по существу не развита. Но вместе с тем, мощного развития достигает мезодермальный компонент, который составляет основу прекрасно развитой сосудистой сети в эмбриональной части плаценты. Таким образом, аллантоис у млекопитающих, сохранив своё участие в газообмене, дополнил «сервисный» набор функцией питания.
Переход к внутриутробному развитию привел к специализации и изменению анатомического и гистологического строения яйцеводов, преобразовавшихся в матку, и к формированию провизорного органа – плаценты, осуществляющего многообразные отношения между зародышем и материнским организмом.
1. Дробление зародыша мыши в яйцеводе (срез). Оплодотворение яйца Млекопитающих, как и у всех Amniota, происходит в воронке яйцевода. Дробление зародыша осуществляется по мере его движения по яйцеводу. На препарате видна стенка яйцевода, выстланного цилиндрическим эпителием, основная толща стенки яйцевода представлена рыхлой соединительной тканью. Дробящийся зародыш, окруженный тонкой яйцевой оболочкой – zona pellucida, находится в просвете яйцевода.
На рисунке отметить: дробящийся зародыш, бластомеры, zona pellucida, стенку яйцевода.
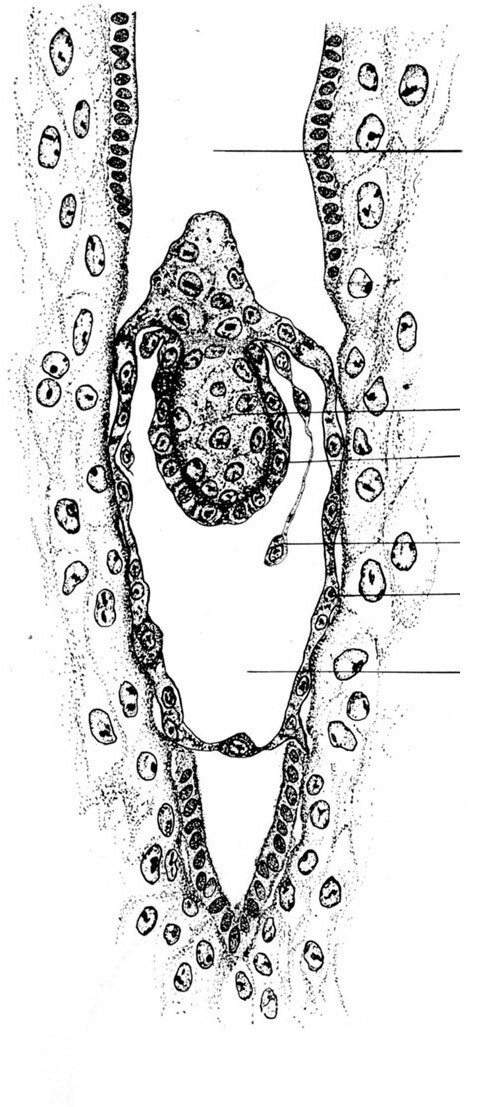
2. Бластоциста крысы в полости матки (срез). В связи с ранней сегрегацией на зародышевый и внезародышевый (провизорный) материал в эмбриогенезе Млекопитающих, развивающуюся структуру от оплодотворения и до окончания имплантации принято называть концептусом. Концептус, начиная со стадии 16 бластомеров, называется морулой, на поверхности которой располагаются уплощённые клетки, соединённые с помощью плотных контактов в единый слой, изолирующий от внешней среды внутренние клетки. Клетки поверхностного слоя (трофобласт, от греч. трофеин – питание, бластос - зародыш) отличаются от внутренних клеток по многим параметрам, в том числе и своей способностью «накачивать» жидкость внутрь зародыша. В результате накопления жидкости зародыш приобретает вид пузырька. К одному из участков стенки пузырька прилежит группа внутренних клеток – внутренняя клеточная масса (ВКМ или эмбриобласт). Участок трофобласта, покрывающий эмбриобласт, называют полярным трофобластом (у мышевидных грызунов он получил название рауберового слоя), трофобласт, окружающий стенку образовавшейся полости, называют муральным трофобластом. Пузырёк с таким строением получил название бластодермического или бластоцисты. Позже за счет ВКМ формируется зародыш и некоторые внезародышевые структуры (гипобласт, амниотическая эктодерма, внезародышевая мезодерма), тогда как трофобласт представляет собой исключительно провизорный участок, в будущем, хорион, образующий зародышевую часть плаценты. После попадания в матку до начала имплантации бластоциста располагается вблизи от стенки полости матки.
На рисунке отметить бластоцисту (эмбриобласт и трофобласт), полость и стенку матки.
3. Строение стенки матки мыши и имплантация зародыша (срез). Стенка матки Млекопитающих состоит из трёх оболочек: 1) тонкой наружной серозной оболочки (serosa); 2) толстой мышечной стенки – миометрия (myometrium) и 3) внутренней слизистой оболочки - эндометрия (endometrium). Серозная оболочка (перитонеальный покров органа) состоит из одного слоя мезотелиальных клеток, под которым лежит тонкий слой рыхлой соединительной ткани. Миометрий состоит из пучков гладких мышечных волокон, разделённых соединительной тканью. Середина миометрия содержит наиболее крупные кровеносные сосуды стенки матки. Слизистая оболочка, выстилающая полость тела и дна матки, состоит из слоя эпителия и собственной пластинки соединительной ткани (lamina propria), связанной с миометрием. Она пронизана простыми трубчатыми железами, которые открываются на поверхности эпителия. У грызунов от просвета в стенку матки отходят тонкие и глубокие дивертикулы – крипты. В период имплантации бластоциста погружается в одну из крипт эндометрия; под влиянием веществ, выделяемых бластоцистой, происходит лизис эпителия крипты и возникает более тесный контакт между трофэктодермой зародыша и эндометрием матки. Имплантация сопровождается глубокими преобразованиями всех слоёв эндометрия матки, который превращается в отпадающую (децидуальную) оболочку.
На рисунке отметить эндометрий, его эпителий и соединительно-тканную часть, миометрий и периметрий: полости матки и имплантационной крипты, имплантирующуюся бластоцисту (в ее составе зародышевый цилиндр, формирующийся за счет эмбриобласта; и эктоплацентарный конус, возникающий за счет размножающихся клеток трофэктодермы, прилежащих к ВКМ); трофэктодерму; продукты лизиса тканей матки.
4. Зародыш мыши в период образования аллантоиса (продольный срез). По завершении имплантации в составе зародышевого цилиндра формируются две полости: амниотическая полость и полость экзоцелома; в составе эктоплацентарного конуса образуется полость ножки плода. Желточная эндодерма (гипобласт), распространяющаяся из области зародышевого щитка, начинает обрастать полость бластоцисты и по завершении этого процесса становится выстилкой полости желточного мешка.
Дно амниотической полости представлено трехслойным зародышевым щитком, который выгнут в сторону желточного мешка. В полость амниона обращена эктодерма, а наружу - в полость желточного мешка – эндодерма, между ними расположен мезодермальный слой, на переднем конце зародыша по медиальной линии располагается утолщенный головной отросток, состоящий из клеток хордомезодермы; на каудальном конце имеется рыхлый мезодермальный вырост, вдающийся в экзоцелом – это аллантоис. Позже произойдет выпячивание формирующегося тела зародыша в полость амниона, после чего топографические отношения зародыша и его провизорных структур станут обычными для Амниот.
На рисунке отметить полости амниона, экзоцелома и полость ножки плода; полость желточного мешка, область формирования тела зародыша, в его составе экто-, мезо - и эндодерму, головной отросток зародыша (нотохорд), аллантоис.
5. ДЕМОНСТРАЦИЯ (бинокуляр). Зародыш кролика на стадии 9 пар сомитов (около 9 суток после оплодотворения). Тоатльный препарат. Зачаток головного мозга представлен тремя мозговыми пузырями. Каждый ряд сомитов латерально окаймлен тяжем мезодермы пронефрического протока. На этой стадии только начинает образовываться головная амниотическая складка, но зачаток аллантоиса уже выступает за пределы тела зародыша.
Зарисовать и отметить: мозговые пузыри, глазной пузырёк, нейропор, нейральную трубку, проамнион, желточные вены, сомиты, сегментационную пластинку, тяжи нефрогенной мезодермы, зародышевую полоску, зачаток аллантоиса.
6. Зародыш кролика в амнионе (поперечный срез). По завершении гаструляции у Млекопитающих, как и у других Амниот, формируется осевой комплекс зачатков с их характерным взаиморасположением. К этому моменту уже вполне сформирован амнион. На препарате отчетливо виден переход эктодермы зародыша в эктодерму амниотической оболочки, а париетального листка спланхнотома – в ее мезодермальный листок. Серозная оболочка и желточный мешок на препарате удалены.
На рисунке отметить элементы осевого комплекса зачатков зародыша, парную спинную аорту, элементы дифференцирующегося нефротома, париетальный листок спланхнотома; эктодермальный и мезодермальный слои амниотической оболочки, полость амниона.
7. Гемохориальная лабиринтная плацента крысы (срез). Гемохориальные плаценты в ходе плацентации зародыша характеризуются разрушением эпителия и соединительнотканной части эндометрия матки, а также стенок сосудов в эндометрии, что приводит к установлению непосредственного контакта между хорионом плода и кровью матери. В гемохориальных плацентах лабиринтного типа этот контакт осуществляется в результате появления в соединительнотканной части хориона системы лакун, по которой курсирует как кровь зародыша (область плацентарного лабиринта хориона), так и кровь матери (область губчатого слоя хориона). Кровь матери изливается в лакуны хориона благодаря разрушению стенок сосудов в децидуальной (прилегающей к хориону) части эндометрия. Следует подчеркнуть, что слияния кровотока матери и зародыша в плацентах никогда не происходит.
На рисунке отметить: материнскую часть плаценты, ее децидуальную область, гигантские клетки трофобласта на границе материнской и фетальной частей плаценты, губчатый слой хориона (в его лакунах кровь матери с безъядерными эритроцитами), плацентарный лабиринт – часть хориона, по которой осуществляется ток крови зародыша, отличающейся от материнской ядерными эритроцитами.
Автор –
