

Для существования каждого вида животных необходимо разрешение трех основных задач: питания, размножения и защиты от опасностей для сохранения особей и вида в условиях борьбы за существование. Движение у позвоночных, и в частности у птиц, является одним из наиболее существенных элементов защиты животного. Рассмотрев связанные с ним стороны биологии птиц, перейдем к рассмотрению их особенностей, связанных с питанием. Условия питания в значительной степени определяют ход жизненных явлений у птиц. Они влияют на географическое распределение птиц, на сезонные перемещения, на темпы размножения и смертности, на условия внутривидовой и межвидовой конкуренции. Необходимость питаться определенным видом пищи обусловливает кормовые стации каждого вида. Сезонные изменения окружающей обстановки частично вызывают изменения условий питания, частично меняют норму потребности организма в пище (в холодное время года при большой потере организмом тепла требуется больше пищи). Кочевки и миграции птиц также находятся в известной связи с условиями питания.
Кормовой режим отдельных видов, весьма различен. Он меняется и по сезонам, и с возрастом птицы. Некоторые виды в отношении питания узкоспециализированы (стенофаги), другие не проявляют предпочтения определенному роду пищи (эврифаги). Питаются птицы и растительной и животной пищей, причем последняя, в общем, преобладает. Остановимся на важнейших особенностях строения птиц, связанных с условиями и способами питания. За относительно немногочисленными исключениями (в частности, к ним относятся совы и хищные птицы), птицы берут пищу клювом. Форма клюва, поэтому весьма разнообразие Длинные клювы имеют птицы, которые достают пищу из воды или с земли (аисты, цапли, кулики и др.). У этих птиц наблюдается корреляция между длиной клюва и длиной ног и шеи. Это обычно неплавающие формы. С другой стороны, длинный клюв характерен для некоторых птиц тропических лесов, питающихся плодами древесных растений, — туканов и птиц-носорогов. Большие размеры клюва у этих птиц компенсируются сильно развитой пневматичностью черепа. Наконец, длинный клюв встречается у многих сосущих цветочный нектар видов (многие колибри, медососы и др.) или у птиц, отыскивающих пищу в складках и углублениях камней или коры (пищухи, стенолазы). У птиц, клюв которых служит для удержания живой и иногда крупной добычи, он умеренной длины или даже короткий, но снабжен крутым крючком на конце верхней челюсти (бакланы, совы, дневные хищники), а иногда и зубцом (сокола). У хватающих крупную добыч птиц нижняя челюсть обыкновенно бывает крупной и высокой (цапли, аисты, чистки, чайки); но иногда и у птиц, питавшихся позвоночными, нижняя челюсть бывает небольшой, короткой и низко (хищные, совы), в последнем случае хватание добычи производится обычно силы вооруженными лапами. У птиц, хватающих на лету насекомых,— ласточек, стрижей, мухоловок — клюв не длинен, но широк и как бы приплюснут, а разрез рта идет далеко назад. У них, как и у других насекомоядных птиц, края рта несут твердые щетинки, облегчающие лов насекомых. У дятлов, долбящих дерево, клюв очень крепкий, прямой и долотообразный; действие его дополняется длинным языком, конец которого усажен острыми шипообразными выступами, крепко придерживающими насекомое. У клестов, вылущивающих из шишек хвойные семена, челюсти крестообразно перекрещиваются и образуют рычаг для поднятия чешуек шишек. У зерноядных воробьиных (вьюрковые и др.) клюв короткий, крепкий, широкий и высокий; нёбная поверхность несет у них резкие борозды и валики; все это является приспособлением для раскусывания и размельчения семян и косточек плодов.
Зубов у современных птиц нет. Среди раннетретичных видов встречались еще формы, имеющие зубы, но, по крайней мере, со среднего эоцена зубатых птиц более уже не встречалось. Размельчение пищи производится у птиц или клювом (например, у хищников), или комбинированными движениями клюва и языка (у зерноядных), или желудком. Добыча часто придерживается одной или двумя ногами. Дятлы и поползни размельчают предметы питания (шишки, желуди и т. п.), защемляя их в дереве (так называемые «дятловые кузницы»). Вороны, чайки и, быть может, бородач раздробляют твердую добычу (раки, раковины, кости и т. п.), бросая ее с высоты на землю. От разнообразия способов размельчения и предварительной обработки пищи зависит и разнообразие устройства и функций языка у птиц. У многих форм язык рудиментарен и служит только для изоляции воздухопроводящих путей; таков язык бакланов, пеликанов, олуш, зимородков, птиц-носорогов, удодов, страусов и некоторых буревестников. Впрочем, такую же функцию выполняет язык и у других видов (механизм таков: когда птица держит в клюве пищу, то конец языка упирается в лежащую посередине нёба щель и дает возможность пользоваться для дыхания носовой полостью). У других птиц язык служит «щупом» (дятлы, поползни), всасывающим насосом (колибри, медососы, нектарницы), хватательным органом (попугаи), тормозом для удержания скользкой добычи (пингвины), теркой (хищные птицы), наконец, сложноустроенным ситом (фламинго, утки, гуси). Вкусовых почек у птиц на языке нет — они расположены на нёбе под языком и в глотке. Чувство вкуса у птиц развито довольно значительно: птицы различают сладкое, соленое, кислое, а некоторые виды птиц и горькое. Слюнные и слизистые железы в полости рта у птиц развиты относительно слабо; они вовсе отсутствуют у веслоногих птиц, глотающих захватываемую в воде скользкую добычу. Характерна яркая окраска рта птенцов — красная, желтая, оранжевая. Кроме того, во рту птенцов часто бывают особые ярко окрашенные выросты. Эта окраска служит для родителей возбудителем рефлекса кормления: яркий, широко открытый рот птенцов побуждает родителей к кормлению (птенцов с закрытым ртом птицы — тут имеются в виду птенцовые — не кормят). Окраска рта, птенца и упомянутые здесь выросты служат родителям и для опознавания своего потомства. Заслуживает внимания тот факт, что у некоторых африканских ткачиков, подкладывающих свои яйца в гнезда других видов, т. е. являющихся так называемыми гнездовыми паразитами, окраска ротовой полости их птенцов сходна с окраской полости птенцов птиц-хозяев.
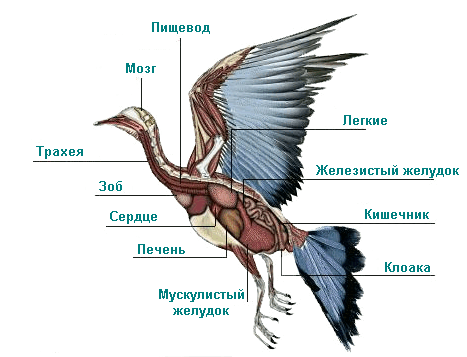
Пищевод птиц довольно растяжим, в особенности у заглатывающих крупную добычу видов (пеликаны, чайки, цапли, бакланы); характерно и часто встречающееся образование, так называемый зоб — богатое железами расширение пищевода. У тех птиц, которые сразу поглощают большое количество пищи, но подолгу иногда голодают, зоб, служит резервуаром для пищи, постепенно поступающей в желудок. У других, например у куриных, попугаев, уже в зобе начинается предварительная обработка пищи. У хищников в зобе накопляются непереваренные части пищу — кости, шерсть, перья и т. п. Передний отдел желудка птиц — так называемый железистый желудок — выполняет функции химической обработки поступающей пищи, а задний — мускульный желудок — перерабатывает пищу механически. Задний (нижний) конец желудка отделяется от кишечника кольцеобразным мускулом-сжимателем (сфинктером), препятствующим, проникновению в тонкие кишки обломков костей и других твердых или острых частей пищи. У рыбоядных видов птиц (цапли, бакланы, поганки, пингвины) и у некоторых других в заднем конце желудка имеется еще третий отдел — так называемый пилорический мешок; функция его — продление нахождения в желудке пищи для лучшей ее обработки. Железистый желудок более всего развит у птиц, сразу заглатывающих больше количество пищи (рыбоядных и хищных). Секрет пищеварительных желез у птиц действует весьма энергично: у марабу и многих хищных он полностью или в значительной мере растворяет кости, а у бакланов, цапель и уток — рыбью чешую, у сов и сорокопутов кости вовсе не перевариваются. Для всех видов птиц непереваримы хитин, кератин и клетчатка (последняя, быть может, у куриных, утиных и голубей частично усваивается вследствие деятельности кишечных бактерий). Мускульный желудок у некоторых птиц отличается сильным развитием мускулатуры, образующей к тому же сухожильные диски. Стенки желудка работают в этом случае как жернова и перетирают твердую и грубую пищу. Так устроен мускульный желудок у зерноядных и питающихся твердыми членистоногими моллюсками птиц (куриные, гусеобразные, страусы, журавли, многие воробьиные, многие голуби). У других птиц мускулатура в мускульном желудке развита незначительно, и в нем продолжается главным образом химическая обработка пищи ферментами, стекающими из железистого желудка. Так устроен мускульный желудок у мясоядных, рыбоядных и плодоядных птиц. У многих видов птиц трубчатые железы мускульного желудка выделяют секрет, образующий затем периодически сменяющуюся твердую кератиновую оболочку, так называемую кутикулу. Это также аппарат для перетирания пищи. Наконец, у многих птиц механическое воздействие мускульного желудка на пищу усиливается еще и тем, что они заглатывают песок, камешки или твердые семена растений. Переваренная пища поступает из желудка в кишечник, сначала в двенадцатиперстную кишку, затем в тонкие кишки. У большинства птиц имеются слепые кишки. Иногда они несут пищеварительные функции иногда являются вместе с тем и лимфатически-эпителиальным органом, иногда только последним; у некоторых видов слепые кишки рудиментарны или даже вовсе отсутствуют. Наибольшего развития достигают они у растительноядных птиц (впрочем, бывают и, исключения). Прямая кишка у птиц служит для скопления непереваренных остатков пищи; конец ее переходит в клоаку — орган, общий для птиц и пресмыкающихся. В клоаку открываются также протоки мочевой и половой систем, и в ней на спинной стороне находится так называемая фабрициева сумка, подвергающаяся у взрослых птиц (в возрасте 8—9 месяцев) редукции, но хорошо развитая у молодых. Функция этой сумки заключается в образовании лимфатических клеток и оксифильных лейкоцитов. Печень у птиц относительно очень велика, желчные протоки ее впадают в двенадцатиперстную кишку. У большинства видов имеется желчный пузырь, что связано с необходимостью одновременного снабжения кишечника большим количеством желчи (для обработки водянистой и жирной пищи). Поджелудочная железа у птиц имеет довольно разнообразную форму, но всегда хорошо развита и относительно больше, чем у млекопитающих. Размеры ее и значение обратно пропорциональны желчному пузырю: крупнее всего, она у зерноядных, мельче у мясоядных птиц. Относительный общий оборот энергии у птиц очень высок, особенно у мелких воробьиных, у крупных же видов он приближается к величине оборота энергии млекопитающих. У серой вороны, например, при окружающей температуре 20—22°С общий оборот энергии составляет 840 кал на 1 м2 поверхности тела в сутки, у сарыча — 780 кал, у курицы (при температуре 23°С) — 580 кал; в то же время при нейтральной температуре (32—36° С), т. е. при минимальной теплоотдаче, оборот энергии у щегла составляет 1534 кал, у серого сорокопута даже 1775 кал на 1 м2 поверхности в сутки. Оборот энергии и потребность в питательных веществах, а в соответствии с этим и сердечная деятельность, и работа дыхательного аппарата меняются, в зависимости от внешних условий и периодических изменений внутреннего состояния организма. У самцов расход энергии повышается в период спаривания, у самок — в период откладки яиц. Повышение расхода энергии связано с периодом линьки. Понижение оборота энергии наблюдается у насиживающих птиц, что можно, рассматривать как приспособление к долгому и неподвижному нахождению на гнезде. Понижение, внешней температуры ниже известных пределов вызывает повышение расхода энергии на поддержание температуры тела. Например, падение внешней температуры с 32,6 до 9,8° вызывает у воробья повышение расхода кислорода в три раза. Небольшие птицы для сохранения тепла вынуждены расходовать больше энергии, чем крупные (размеры поверхности тела растут в квадрате, а объем в кубе, следовательно, у крупных птиц отношение поверхности тела к объему выгоднее). Мелкие птицы при значительном понижении температуры расходуют на терморегуляцию организма более половины получаемой от питания энергии. Зимой для птиц вследствие похолодания и сокращения дня наступают критические моменты, и при сильном понижении температуры может наступить смерть от истощения: наступление темноты прекращает возможность кормления, и птица не может получить достаточных источников энергии.
Существенное значение для терморегуляции птиц имеет оперение и его сезонные изменения. При линьке осенью у многих видов отмечается увеличение пуховой части пера или (при двойной линьке в год) увеличение количества перьев по сравнению с теплым временем года. Географические формы (подвиды), живущие на севере, отличаются от своих южных родичей более густым и пышным оперением (трехпалые дятлы, большие пестрые дятлы, гаички, кречеты). Большое значение для северных птиц имеет белая окраска их оперения, при которой в пере образуются пузырьки воздуха, создающие теплоизолирующий слой. Значение пера для сохранения тепла ясно само по себе, но конкретное представление об этом лучше всего видно из опыта Джиаиа (1929): у большого серого сорокопута при понижении температуры с 28 до 0,6° расход энергии возрос на 50%, но когда птица была ощипана, то такая же разница в температуре вызвала увеличение расхода энергии в три раза, т. е. на 200%. Другие приспособления к холодной температуре: отложение подкожного жира (особенно у водных птиц), работа воздушных мешков (сохраняющих в себе согретый воздух), некоторое увеличение размеров птиц у северных форм одного и того же вида по сравнению с южными, наконец, относительное увеличение размеров сердца. Голодание вызывает у птиц понижение температуры. Вообще же говоря, у тех видов, которые имеют более высокую температуру тела и высокую потребность в кислороде и более подвижны, потребность в пище выше и усвоение ее идет быстрее. Противоположные показатели указывают на меньшую потребность в пище. Поэтому, например, птенцы певчих птиц погибают уже через несколько часов после начала голодания, тогда как крупные виды могут прожить без еды около месяца (белая сова — 24 дня, орлан-белохвост — 45 дней, беркут — 21 день, домашние куры — 26—31 день). Потеря веса при этом может достигать 30—40%. Потребность организма птиц в воде относительно невелика. Объясняется это незначительностью кожного испарения, а также тем, что из мочи вода всасывается организмом птицы обратно во время нахождения мочи в верхнем отделе клоаки. Многие мясоядные и плодоядные виды поэтому вовсе не пьют. Пищеварительный процесс проходит у птиц очень быстро и энергично. При этом быстрее перевариваются и усваиваются мясо и плоды, медленнее — семена. За сутки птица может съесть очень много, и максимум в этом случае часто сильно превышает необходимый минимум. Мелкие совы (домовые сычи) переваривают мышь за 4 часа, серый сорокопут — за 3 часа; водянистые ягоды у воробьиных проходят через кишечник за 8—10 минут, зерна у курицы — за 12—24 часа. Насекомоядные птицы наполняют свой желудок пять-шесть раз в день, зерноядные — дважды. Один-два раза в день едят хищники. Мелкие птицы съедают за сутки сухих пищевых веществ около 1/4 своего веса, крупные — значительно меньше (около 1/10). Птенцы едят больше. Точными наблюдениями установлено, что ласточки, синицы, скворцы и другие мелкие птицы во время выкармливания птенцов подлетают к гнезду с пищей сотни раз в сутки. Так, большая синица приносит пищу 350—390 раз, поползень — 370— 380 раз, горихвостка — 220—240 раз, большой пестрый дятел — 300 раз, а американский крапивник даже 600 раз. При этом прирост веса у птенцов в сутки составляет 20—60% первоначального веса. За первые семь-восемь дней вес у птенцов воробьиных увеличивается в 5—6 раз. Понятно поэтому, что птенец за сутки съедает больше пищи, чем весит сам. Это обстоятельство определяет громадное значение насекомоядных птиц в жизни природы и в хозяйстве человека. При большой интенсивности роста птиц и довольно значительном числе яиц в кладках (которых к тому же у многих видов бывает нормально по две в год, а у некоторых и по три) одной паре воробьиных птиц приходится в среднем выкармливать 10—15 молодых ежегодно.
Наконец, сравнительно недавно установлено еще одно замечательное биологическое свойство птиц: обилие пищи и благоприятные кормовые условия вызывают у них усиленное размножение. Таким образом, у многих видов в годы, благоприятные по условиям питания, число яиц в кладке больше, чем в менее благоприятные годы. Иногда в «урожайные» кормами годы у птиц появляются и дополнительные кладки. Наоборот, в неблагоприятные по кормовым условиям годы интенсивность размножения уменьшается (число яиц в кладке меньше), а смертность среди молодых птиц становится очень высокой. Заслуживает внимания еще одна черта. При обилии пищи птицы едят больше. Например, по сделанным в Западной Европе наблюдениям, в «мышиные» годы один сарыч съедает ежедневно до 14 мышей и полевок, а в обычные средние годы — до 5 штук, пустельга ест соответственно 9 и 2 мыши, ушастые совы — 12 и 4 и т. д. Следует учесть, что одна полевка, по подсчетам наших экологов, уничтожает до 2 кг зерна в год. Наконец, обильное появление какого-либо рода пищи приводит иногда к тому, что ею начинают питаться те виды птиц, которые обычно этим родом пищи пренебрегают. Интересны результаты наблюдений , сделанных им в 1936 году в Северо-Западном Казахстане: при появлении большого количества саранчи ею стали питаться даже утки. Можно, таким образом, говорить о том, что кормовые условия определяют многие стороны жизни птиц, причем в случае массового размножения того или иного объекта питания птиц он привлекает к себе особое с их стороны внимание. Происходит, следовательно, известного рода естественное регулирование числа массово размножившегося животного. Хорошо известно, что появление где-либо в большом количестве вредных насекомых обычно привлекает птиц. В таких случаях полезность насекомоядных птиц выступает особенно наглядно. Когда, например, в 1893—1895 гг. в Поволжье сильно размножился вредитель лесов — непарный шелкопряд, то местные наблюдатели отмечали необычайный налет кукушек. Размножение вредителей полеводства — щелкунов привлекает грачей, которые выкапывают из земли и поедают личинок этих жуков, так называемых проволочных червей. По некоторым подсчетам, грач съедает более 8000 проволочных червей в год. Имеются наблюдения, как грачиная стая за один день полностью очистила от этих, вредителей площадь в 6 га. Размножение саранчи вызывает усиленное размножение и скопление разных скворцов, в частности розовых. За странствующей саранчой следуют самые разнообразные виды птиц. Размножение мышей вызывает усиленную деятельность на полях хищных птиц — сов, сарычей, мелких соколов. За странствующими леммингами в тундрах и лесотундрах следуют многочисленные белые совы, большие чайки и поморники, мохноногие канюки и даже сапсаны. Пища многих видов птиц состоит из животных, имеющих для человеческого хозяйства отрицательное значение. Это насекомые и мелкие млекопитающие, в первую очередь грызуны. Размножение и тех и других идет и может идти очень быстро. И вот в борьбе с этими вредителями заключается основное положительное значение птиц для хозяйства. Промыслово-охотничьи и домашние птицы приносят прямую пользу человеку, но значение ее, в противовес широко распространенному еще и теперь мнению, невелико по сравнению с той пользой, которую птицы приносят истреблением полевок, мышей, вредных насекомых, их яиц и личинок. Нет сомнения, что с хозяйственной точки зрения именно эта сторона деятельности птиц представляется наиболее важной и значительной. Значение опасности для сельского хозяйства со стороны вредителей никоим образом не следует преуменьшать. Если в наше время — время высокой техники — они не могут довести положение до катастрофы, то все же причиняют весьма тяжкий ущерб. В дореволюционной России убытки полеводства от вредителей определялись (конечно, с известным приближением) в 900 млн. руб. в год, убытки лесоводства — в 300 млн. руб., убытки садоводства и огородничества—в 90 млн. руб. В Соединенных Штатах Америки убытки сельского хозяйства от животных-вредителей в 1921 году определялись в биллион долларов, а польза от истребления насекомых птицами — 444 миллиона долларов; следовательно, птицы уменьшили ущерб более чем на одну треть в относительных цифрах и на громадную абсолютную величину. Все эти расчеты, конечно, приблизительные, но они дают представление о масштабе и общем значении этого явления.
Существенно еще одно соображение. Из числа известных видов птиц огромное большинство относится к отряду воробьиных, объединяющему в себе, за редкими исключениями, насекомоядных птиц, или птиц, выкармливающих птенцов насекомыми. Кроме того, количество особей этих мелких и средних видов неизмеримо больше, чем число особей крупных видов, так что не будет преувеличением считать, что насекомоядные птицы составляют около 90% всего числа особей живущих в настоящее время птиц. Если так, то можно, пожалуй, согласиться с одним американским автором, высказавшим мысль, что «если бы все птицы были уничтожены, то сельское хозяйство в Соединенных Штатах стало бы невозможным». Не следует представлять себе дело, таким образом, будто бы птицы сами по себе могут уничтожить вредителей при их массовом размножении, но роль их в истреблении грызунов и насекомых в «нормальные» годы очень велика и может быть охарактеризована как «контроль» над размножением вредителей, как весьма существенное орудие удержания числа вредителей на низком уровне. Другие стороны деятельности птиц, связанные с питанием, также небезразличны для человека. Многие зерноядные птицы способствуют распространению семян (последние иногда сохраняют всхожесть и после прохождения через кишечник птицы), в южных странах многие виды активно способствуют опылению растений. Хищные птицы, охотящиеся за другими птицами и зверями, играют известную положительную роль в качестве орудия отбора. Известное число хищников способствует поддержанию здоровья видов, являющихся их добычей, так как в первую очередь ими добываются больные или слабые экземпляры. Птицы, питающиеся падалью, приносят известную пользу в санитарном отношении.
Отрицательные с точки зрения хозяйственных интересов человека черты птиц, связаннее с питанием, могут быть сведены в общих чертах к истреблению полезных диких животных и дикорастущих растений, к конкуренции с более полезными видами животных, к причинению вреда культурным растениям, к поеданию домашних, животных. При этом следует иметь в виду, что у нас нет никаких оснований говорить, что та или иная, птица: должна считаться абсолютно полезной или абсолютно вредной. Ни пользы, ни вреда «вообще» птицы не приносят. Поэтому не может ставиться вопрос об абсолютной охране или абсолютном уничтожении какого-либо вида птиц. И полезной и вредной птица, как и другое животное, может быть только в определенных условиях и в определенное время. Меняется обстановка — меняется и хозяйственное значение птиц. Скворцы, например, приносящие весной и летом пользу истреблением насекомых, в некоторых районах на пролете и зимовках могут быть определенно вредными для садов, и еще совсем недавно в Тунисе борьбу со скворцами вели массовым применением взрывчатых веществ. Вороны вредят разорением гнезд полезных птиц, в частности водоплавающих, но вместе с тем истребляют насекомых, мышей и полевок. Большой пестрый дятел питается вредными для леса насекомыми, но вместе с тем уничтожает известное число семян деревьев, а иногда вредит и самим деревьям (так что иногда и местами, как, например в Бузулукском бору, вреда от большого пестрого дятла, мешающего нормальному возобновлению сосны, больше, чем пользы). Воробей ест ягоды, вытесняет с мест гнездовья полезных насекомоядных птиц, но и сам выкармливает птенцов насекомыми. Сокол-сапсан питается водоплавающими и другими полезными птицами, но вместе с тем в тундре вблизи его гнезд песцы оставляют в покое гнезда других птиц, так как сокол энергично нападает на песцов и прогоняет их из окрестностей своего гнезда, оказывая этим существенную помощь всему окрестному птичьему населению. Ястреб-тетеревятник питается полезными птицами, но способствует естественному отбору и местами справедливо ценится в качестве прекрасной ловчей птицы. Мы не коснемся здесь весьма немаловажного вопроса об эстетическом значении птиц. Полезно подчеркнуть, что в фауне России, насчитывающей более 750 видов птиц, существенно вредными является, во всяком случае, менее десятка видов. Заимствованное у западноевропейских владельцев охотничьих угодий и их егерей и, к сожалению, прочно укоренившееся и широко распространенное мнение о «вреде» хищных птиц должно быть решительно, отброшено. Громадное большинство хищников приносит пользу истреблением грызунов и насекомых; другие, например крупные сокола — сапсаны, кречеты, хотя и охотятся преимущественно на птиц, но являются редкими, да к тому же в сколько-нибудь заметном числе живут в таких районах (север), где дичные богатства еще недостаточно использованы человеком. Конкурентами последнего они никоим образом не являются, а вместе с тем служат одним из лучших украшений нашей природы; да и размножение хищных птиц идет сравнительно медленным темпом. Это не значит, что не следует бороться с хищниками, повадившимися ловить голубей, домашнюю птицу, или с ястребом, разгоняющим тока тетеревов в организованном охотничьем хозяйстве, и т, п.
Кормовые условия отражаются на географическом и на стационарном распределении птиц. В особенности это относится к тем видам, которые являются стенофагами, т. е. узкоспециализированы в питании. Африканский грифовый орлан встречается только там, где растет тот вид пальмы, плодами которой он питается. Многие птицы, питающиеся определенными растениями или у которых определенный вид растения, преобладает в кормовом рационе, встречаются только там, где имеются эти растения. Так, например, шотландский граус тесно связан в своем распространении с багульником, клесты— с определенными видами хвойных деревьев, медососы, колибри и др.— c наличием тех растений, нектаром которых они питаются. Всеядных птиц, в сущности, немного: примером их могут служить врановые. В общем же для каждого вида птиц свойственна определенная специализация как в выборе пищи, так и в способах ее добывания. К сожалению, вопросы эти изучены еще недостаточно. Между тем некоторые определенные вещества, поглощаемые птицами хотя бы в небольшом количестве и изредка, имеют, по-видимому, очень большое значение для нормального функционирования организма птицы. Например, у молодых хищных птиц, не получающих костей, развивается рахит и нарушается нормальный ход линьки. Для тетеревиных необходимо, временами заглатывать хвою, служащую, вероятно, для очищения желудка от глистов. Изменение внешних условий, определяющих условия питания, имеет для птиц большое значение. Эти изменения особенно сказываются в тех районах, где климатические перемены по временам года значительны или где различного рода метеорологические условия (снеговой покров, влажность, температура и т. п.) сильно колеблются. О влиянии температуры на потребность организма птиц в пище и о влиянии света на возможность удовлетворять эту потребность уже сказано выше. Снеговой покров имеет также большое значение для видов, кормящихся на земле. Поэтому, например, многие зерноядные птицы зимуют в Монголии, где зимы очень суровы, но снега мало. С другой стороны, например, в. Лапландии за полярным кругом можно и зимой встретить довольно разнообразный состав мелких воробьиных: гаичек, большую синицу, пищуху и др. Эти птицы добывают пищу с деревьев и менее зависят от снежного покрова. По той же причине птицы, добывающие пищу из щелей и других укрытий или на вертикальных стволах деревьев в коре и т. п., например крапивники, поползни и упомянутые уже пищухи, не отлетают на зиму, а остаются в холодной и умеренной зонах на родине. Даже в условиях арктической полярной ночи зимуют птицы, лишь бы была у них возможность добывать себе пищу. Например, у берегов Гренландии полярный чистик зимует у полыней и разводий под 77° и даже 78°30' северной широты, у Шпицбергена — даже под 80° северной широты. В тропиках и субтропиках основной климатической причиной изменения условий питания птиц служит наступление засушливого времени гада. Исчезновение насекомых, уменьшение количества насекомых, периодические изменения в жизни растений — все эти факторы определяют пищевой режим птиц и соответственно влияют на их распространение. Если у одних видов эти перемены вызывают передвижения, то у других с ними связаны сезонные изменения пищевого режима. Белые куропатки, например, летом питаются преимущественно ягодами и насекомыми, осенью ягодами, а зимой побегами ивы. Ворон в Северной Сибири летом всеяден, зимой питается преимущественно пеструшками. Скворцы летом питаются главным образом насекомыми, осенью и на зимовках, кроме того, плодами и ягодами. Таких примеров можно было бы привести множество. Урожай и неурожай кормов чрезвычайно влияют на жизнь птиц. Периодические количественные колебания в животном населении и в растительном покрове вызывают периодические колебания условий существования птиц, для которых те или иные животные и растения служат пищей. К этим явлениям относится урожай и неурожай плодов и ягод, обилие или малочисленность насекомых, массовое размножение или вымирание грызунов и т. п. Массовое появление объектов питания вызывает и массовое появление соответствующих видов птиц, и наоборот. Например, при неурожае рябины из Северной Европы массами откочевывают свиристели, при неурожае шишек — клесты, ореховки и др. Более или менее длительные изменения условий питания вызывают иногда и изменения границ области распространения. Так, домовый воробей постепенно расселялся, следуя за человеком, но замена лошадей автомобилями вызвала уменьшение числа воробьев у северной границы его распространения — в Скандинавии и сильное сокращение его численности в североамериканских городах. О влиянии условий питания на размножение и смертность уже говорилось. Здесь приведем только некоторые цифры. В Лапландии в «лемминговые» годы у ястребиной совы бывает 11—13 яиц, у бородатой неясыти — 7—9 яиц, у филина — до 6, у ушастой совы — 7—9, у белой совы — 11—12. Даже у лапландского кречета в исключительно обильный леммингами год близ города Каутокейно на северо-востоке Норвегии находили кладки по 7—9 яиц. О вторых кладках в богатые кормами годы у тех видов, у которых обычно бывает только одна кладка, уже говорилось. С другой стороны, в годы неурожайные, при уменьшении числа грызунов, питающиеся ими хищники имеют меньшее число яиц в кладках, а смертность среди птенцов выше. По-видимому, следствием плохих кормовых условий можно объяснить явление каннибализма среди птенцов многих видов хищных — ястребов, орлов и других птиц, когда младший из птенцов становится жертвой старших. Особенно заметно влияние условий питания на размножение птиц на севере, где в связи с этим наблюдаются периодические негнездования. Такие колебания численности и «отказы» от гнездования установлены в Арктике для хищных птиц и некоторых водоплавающих, а в других широтах — для многих куриных (рябчики, куропатки, перепела, фазаны и др.). Условия питания, несомненно, лежат в основе возникновения птичьих перелетов, хотя, конечно, современная картина этого явления весьма сложна и определяется, по-видимому, целой совокупностью внешних и внутренних причин. К вопросу о перелетах мы вернемся ниже.
Цикл явлений в жизни птиц, связанных с размножением. Для половой системы птиц характерно то обстоятельство, что период ее активности у громадного большинства видов ограничен строго определенным временем в году, причем в состоянии покоя размеры половых желез буквально в десятки раз меньше, чем в период активности. В строении половой системы самок характерна ее асимметрия: правый яичник, как правило, отсутствует, правый яйцевод отсутствует всегда. В период размножения объем яичника весьма возрастает, и так как яйца в нем находятся на разных стадиях развития, то весь орган принимает как бы гроздевидную форму. По окончании откладки яиц яичник быстро уменьшается, и размеры его доходят до размеров яичника периода покоя еще в то время, когда птица насиживает. Точно так же в связи с наступлением сезона размножения увеличивается в объеме и яйцевод. Например, у домашней курицы яйцевод в период покоя имеет около 180 мм в длину и 1,5 мм в просвете, в период кладки — около 800 мм в длину и около 10 мм в просвете. Все отделы яйцевода в это время становятся более обособленными, чем в другое время года. После периода кладки яйцевод спадается, канальцы его желез редуцируются, просвет его остается неровным и местами расширенным. У птицы же, не несшей яиц, яйцевод имеет вид ровного и на всем протяжении тонкого канальца. Эти различия в состоянии яйцевода могут служить надежным признаком при определении возраста у осенних и весенних птиц. Весьма характерным приспособлением к выводу потомства у птиц служит развитие так называемых наседных (высиживательных) пятен. Наличие этих пятен облегчает обогревание кладки. Кожа в области наседных пятен отличается особой рыхлостью соединительной ткани; жировой слой тут обычно пропадает; пух, а иногда и перья и их зачатки выпадают; кожные мускульные волокна редуцируются; одновременно усиливается питание этих мест кровью. Вполне развитое наседное пятно представляет собой участок голой и слегка воспаленной кожи. Для каждого вида птиц характерно определенное расположение наседных пятен; они бывают то парными, то непарными. У воробьиных, буревестников, чистиков имеется одно пятно, у фазановых, куликов, чаек, хищных — два брюшных и одно грудное. Величина наседных пятен находится в известном соответствии с величиной кладки. Гуси и утки наседных пятен не имеют; у них, однако, в период кладки яиц развивается особый длинный пух, который выдергивается птицей; этим пухом насиживающая птица окружает яйца в гнезде, и он служит отличным средством для защиты их от охлаждения. Олуши не имеют наседных пятен, но обогревают яйца, прикрывая их сверху своими перепончатыми лапами; чистики и пингвины подкладывают лапы под яйца. У этих птиц, по-видимому, в лапах имеются особые артерио-венозные анастомозы, обеспечивающие усиленное питание кровью этих частей тела. У пингвинов сверх того имеется особый кожистый выступ, или карман, вблизи клоаки, произвольно растяжимый и позволяющий насиживающей птице прикрывать яйцо кожей. Кроме только что указанных изменений в организме птиц в связи с периодом размножения, имеются и другие, в частности у многих видов развивается яркий брачный наряд. Различие по внешнему виду между самцами и самками обозначается как половой диморфизм. Внешние признаки полового диморфизма не могут быть уложены в какую-либо общую схему. Пингвины, буревестники, веслоногие, поганки, гагары, вертишейки, стрижи, многие щурки и зимородки не имеют различий между полами ни в окраске, ни в размерах. Самцы и самки мелких воробьиных, большинства хищных, сов, куликов, чаек, чистиков, пастушков и других птиц отличаются только размерами. У других видов самцы более или менее резко отличаются от самок по окраске. Обычно окраска самца ярче у тех видов, у которых самец не принимает участия в заботах о потомстве. В этих случаях (утки, многие куриные) самки часто имеют резко выраженную покровительственную окраску. У тех же видов, у которых заботу о потомстве несут самцы (цветные бекасы, плавунчики из куликов, некоторые зимородки, трехперстки и др.), самки несколько ярче, чем самцы. Различия в окраске проявляются обычно после достижения половой зрелости, но иногда и раньше (дятлы, воробьиные и др.). У многих форм, имеющих две линьки в году, диморфизм окраски заметен только в определенное время года, а именно в период размножения. Яркость окраски самцов особенно характерна для северных уток (но не гусей), многих куриных (фазанов, турачей, глухарей, тетеревов), многих воробьиных (так называемых райских птиц, иволг, вьюрков, горихвосток и т. д.). В родственных группах различия окраски полов, в общем, носят сходный характер даже у разных видов (у иволг самцы ярко-желтые или красные, самки тускло-зеленоватые с продольно испещренной брюшной стороной тела; у многих вьюрков в окраске самцов имеются красные цвета, отсутствующие у самок, например у щуров, клестов, снегирей, в особенности у чечевиц и т. д.). Иногда у самок развивается окраска, сходная с окраской самцов (так называемая петухоперость окраски у тетеревиных, у некоторых воробьиных - горихвосток, жуланов и др.). Кроме того, с возрастом у самок с функционирующими половыми железами наблюдается иногда появление особенностей, сходных с окраской самца; это бывает, например, у хищных птиц (дербники и др.). Половые различия в окраске выражаются не только в цвете оперения, но и в цвете других частей организма (клюв, радужина, голые части кожи, даже язык). У кукушек окраска самцов однотипна (серая), самок диморфна (кроме серой окраски, имеется еще рыжая). Половые различия выражаются, кроме того, в наличии выростов и придатков кожи на голове (например, у куриных), в развитии отдельных перьев (хохлы, длинные кроющие хвоста у павлинов, перья на крыле и хвосте у райских птиц, длинные рулевые у фазанов и т. д.), в пропорциях, размерах и форме отдельных частей тела, в устройстве внутренних органов (голосовой аппарат многих видов, горловой мешок самца дрофы и т. п.), в общей величине. У самцов куриных птиц развиваются шпоры на ногах, у самцов и самок многих видов различна величина клюва (у птиц-носорогов, уток, турпанов, некоторых воробьиных и др.). Как правило, самцы бывают крупнее самок. Это особенно выражено, у куриных и дроф. У других групп самки больше самцов. Это наблюдается у тех видов, у которых заботу о потомстве несут самцы (у плавунчиков, цветных бекасов из куликов, трехперсток, тинаму, некоторых кукушек, киви и казуаров). Большая величина самок, однако, встречается и у тех видов, у которых главная часть заботы о потомстве лежит на самках (у большинства дневных хищных, сов, многих куликов).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
